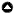|
|
|
Copyright © 2003 by the author(s). Published here under license by The Resilience Alliance.
Go to the pdf version of this article.
The following is the established format for referencing this article:
Tankersley, Jr., R. and K. Orvis. 2003. Modeling the geography of migratory pathways and stopover habitats for neotropical migratory birds. Conservation Ecology 7(1): 7. [online] URL: http://www.consecol.org/vol7/iss1/art7/
A version of this article in which text, figures, tables, and appendices are separate files may be found by following this link.
Report Modeling the Geography of Migratory Pathways and Stopover Habitats for Neotropical Migratory Birds Roger Tankersley, Jr.1 and Kenneth Orvis
1Tennessee Valley Authority
- Abstract
- Introduction
- Methods
- Results
- Discussion
- Conclusion
- Responses to this Article
- Acknowledgments
- Literature Cited
- Appendix 1
- Appendix 2
Intact migratory routes are critical for the stability of forest-dwelling, neotropical, migratory bird populations, and mortality along migratory pathways may be significant. Yet we know almost nothing about the geography of available stopovers or the possible migratory pathways that connect optimal stopovers. We undertake a spatial analysis of stopover habitat availability and then model potential migratory pathways between optimal stopovers in the eastern United States. Using models of fixed orientation and fixed nightly flight distance between stopovers during spring migration, we explore whether a simple endogenous migratory program is sufficient to ensure successful migration across the modern landscape.
Our model runs suggest that the modern distribution of optimum stopovers in the eastern United States can be adequately exploited by birds following migratory pathways defined by fixed-orientation and fixed-distance nightly flights. Longer flight distances may increase the chances of success by enabling migrants to bypass locales offering little habitat. Our results also suggest that most southwest–northeast migratory pathways through the Appalachian mountains are intact. Lack of optimal habitat at key locations in the Southeast causes many modeled pathways to fail. We present a speculative view of regional migration patterns implied by predominant ideas found in stopover ecology literature, and demonstrate the need for broad-scale migration research, in the hope that our approach will foster other continental- and regional-scale projects.
KEY WORDS: Conservation, Forest fragmentation, Habitat, Migration, Modeling, Neotropical migratory birds, Stopover ecology.
Published: April 7, 2003
Breeding Bird Survey (BBS) results suggest a complex geographic pattern of population decline and recovery for forest passerine landbirds (Robbins et al. 1989, Askins et al. 1990, Finch 1991, Brawn and Robinson 1996, Villard and Maurer 1996). Over two-thirds of forest species in eastern North America migrate to wintering grounds in the Caribbean and Central and South America (Keast and Morton 1980), and many western species migrate through eastern North America (Moore et al. 1993). Intact migratory routes are critical for survival (Hutto 2000), and mortality during migration may be significant (Moore and Aborn 2000), yet we know almost nothing about which stopover habitat types are most important or how their distribution and quality define intact migratory pathways (Moore and Simons 1992). Spatial analysis of stopovers and pathways can provide a geographic framework for research in stopover ecology: the study of stopover habitat composition, distribution, and relative importance in life-histories of birds (Moore et al. 1995). Most stopover ecology research has focused on local habitat use (e.g., Hall 1981, Moore and Kerlinger 1987, Winker et al. 1992, Rappole 1995, Yong et al. 1998, Kelly et al. 1999), with practically no research on broad-scale habitat distribution. We examine this geography using simple models of migratory flights to evaluate the basic regional patterns found today and to encourage others to pursue similar broad-scale migratory research.
Migration in birds may be controlled by inherited instincts for habitat preferences (Hutto 2000), migratory duration (Berthold 1996), and migratory orientation (Baker 1984). Although habitat type selection may be fairly inflexible, there is good evidence of flexibility in orientation (Woodrey 2000) and flight distance (Simons et al. 2000). Young birds may follow slightly different pathways than adults, allowing the species to sample many possible migratory routes (Berthold et al. 1992), with successful breeders endowing a predisposition for migration along intact routes (Berthold 1990). If a given pathway is sufficiently degraded, species may adapt through increased genetic representation by survivors following intact pathways (Berthold 1990). Changing patterns of usable pathways may contribute to the geographic pattern of population decline and recovery seen in the BBS (Brawn and Robinson 1996, Villard and Maurer 1996). As an initial step in research on interconnections between the pattern of migration pathways and breeding populations, we examine the geographic dynamics of migration using a model based on habitat, orientation, and flight distance between stopovers.
Migration habitatsMigrant behavior within stopovers suggests birds employ local habitat selection, and the broad-scale distribution of migrants suggests an evaluation of regional patterns (Hutto 2000). Instinctual preferences for broad habitat types (forest, grassland, or wetland) may help migrants identify locations where their feeding and flight morphologies are best suited to exploit habitat structure (Petit 2000, Simons et al. 2000). The fact that forest migrants congregate in some stopovers but not others is well documented; whether this congregation represents habitat selection at multiple scales has yet to be examined in migration research.
Research suggests that migrants select stopovers between midnight and 2 a.m. (Kerlinger and Moore 1989), possibly relying on habitat features detectable from the air in low light (Martin 1990). Migrants may track physiographic features, such as mountain ranges (Bruderer and Jenni 1990), coastlines(Able 1972), and rivers (Bingman et al. 1982), although such behavior is difficult to assess. We hypothesize that regional landcover patterns—such as amount and arrangement of forest, road density, or extent of agriculture—play a similar role in defining the broad habitat differences that forest migrants use to evaluate a region of potential stopovers (Moore and Aborn 2000, Hutto 2000, Petit 2000).
Migratory orientationPredominant migrant navigation theories include: a solar compass for diurnal migrants (Schmidt-Koenig et al. 1991), a celestial compass for nocturnal migrants (Wiltschko and Wiltschko 1991), orientation along the Earth’s magnetic field (Wiltschko and Wiltschko 1988), olfactory navigation (Papi 1991), and navigation by infrasound emitted by large topographic features (Hagstrum 2000). The ability of migrants to navigate long distances along migratory routes is supported by many instances of site fidelity (e.g., Berthold 1993, Schmidt-Koenig et al. 1991, Winker et al. 1991, Nisbet 1969). This ability may be manifest as an inherited orientation preference (Berthold 1991), with random genetic changes causing some individuals to follow novel routes (Woodrey 2000).
Springtime migrants follow a “northward preferred” migratory heading in the neotropic–nearctic migration (Berthold 1996). Most species exhibit clusters of orientation toward known breeding grounds (e.g., Moore 1984, Sandberg et. al. 1988, 1991, Sandberg and Moore 1996, Woodrey 2000), with older individuals exhibiting tighter clusters than young birds (Moore 1984, Woodrey and Moore 1997, Woodrey 2000, Berthold 1996). Clustering may represent previous success along optimal routes, as younger birds following novel orientations perish along sub-optimal routes, focusing the species toward orientations useful for migration.
Nightly migratory flight distancesThe potential length of nightly flights between stopovers is a function of morphological characteristics such as body size, wing shape, and flight musculature (Pennycuick 1989), combined with energetic condition (Weber and Houston 1997a) and modified by external factors such as wind, turbulence, and weather (Richardson 1991). Literature reviewed by Kerlinger and Moore (1989) suggests migrants begin migratory flights shortly after sundown, the number of airborne birds peaks around 22:00, and flights are generally complete by 02:00—a flight time of approximately 5 to 6 hours.
Some studies report flight times and speeds for individual migrants (e.g., Cochran et al. 1967, Cochran and Kjos 1985), but we agree with Weber and Houston (1997a) that there have been too few studies and that flight distances vary considerably between studies and between individuals within studies. Modeled distances are more applicable for studying species-level migration strategies (Hedenstrom and Alerstam 1998). Many models use equations based on physiological parameters (Pennycuick 1989, Berthold 1996, Weber and Houston 1997a, 1997b, Hedenstrom and Alerstam 1998, Alerstam and Hedenstrom 1998) incorporating three components: energetic condition, morphological limits of flight speed, and refueling rates at stopovers.
ApproachOur effort represents a first step toward developing an analytical understanding of sub-continental patterns of migration for northbound forest passerine species (Table 1) through the eastern United States. We use regional patterns of stopover habitat quality, based on field data mapped during an earlier study (Tankersley 2002). We model possible migration routes between optimal stopovers very simply, assuming constant migration orientation and nightly flight distance. This speculative view of migration strategy is based on predominant ideas found in stopover ecology literature. By testing a subset of literature assumptions, we test their implications, allowing us to consider whether our view of how birds migrate enables them to successfully exploit the current geographic distribution of stopovers. Our primary goal is to illustrate a technique for regional-scale analysis that will encourage further broad-scale migration studies.
|
Table 1. Forest-dwelling, long-distance migrant bird species
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
We mapped the quality of different forested stopover landscapes in the eastern United States (Fig. 1) using data from an earlier study (Tankersley 2002) in which citizen scientists helped us survey migrant abundance in 128 forest patches during the 1999 spring migration. We correlated migrant abundance with patch-level habitat variables, and also with landscape-level habitat variables inside a 10-km radius (314 km2) circle centered on each patch (cf. Flather and Sauer 1996). Our findings suggest that migrants favor intact forests with low fragmentation, embedded in a landscape containing relatively little agriculture or human development.
|
Fig. 1. Study Area. Includes A) 12 states and the panhandle of Florida, and B) portions of five North American Bird Conservation Initiative (NABCI) Bird Conservation Regions.
|
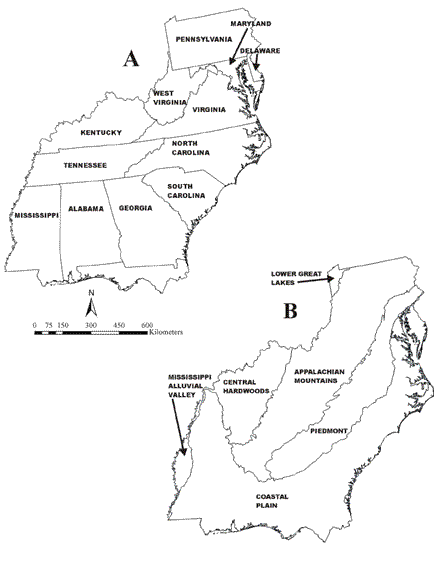
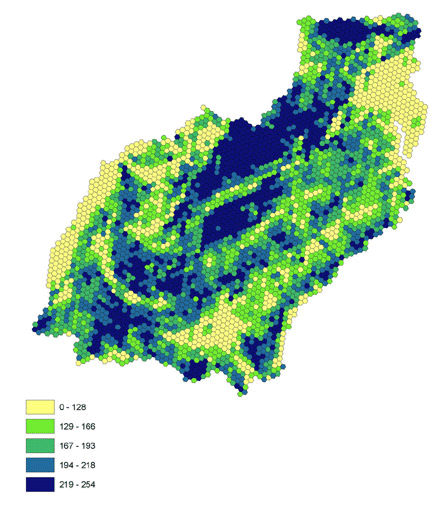
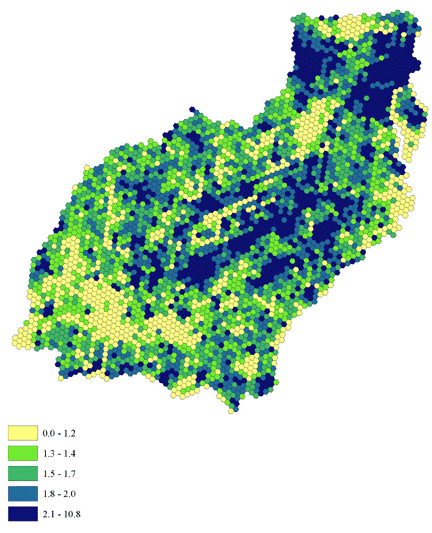
We selected habitat variables based on their correlations with forest migrant abundance and on possibility of aerial evaluation of the variable by migrants: local forest density (Fig. 2), measured using a 65-ha sliding window algorithm (see Appendix 1), percent agriculture (crops and pasture, Fig. 3), and road density (Fig. 4) measured as kilometers of road per square kilometer (ESRI 1997). We approximated a continuous landscape of circles by using hexagons. Local forest density and percent agriculture are related, but we found a strong negative relationship to percent agriculture that was not explained by a simple lack of forest (Tankersley 2002).
|
Fig. 2. Local forest density (quintiles). Darker colors indicate higher density. Data range is 0 255.
|
|
Fig. 3. Percent agriculture (quintiles). Lighter colors indicate more agriculture.
|
|
Fig. 4. Road density (quintiles). Measured as linear road distance (km) per square kilometer within hexagon. Darker colors indicate more roads.
|
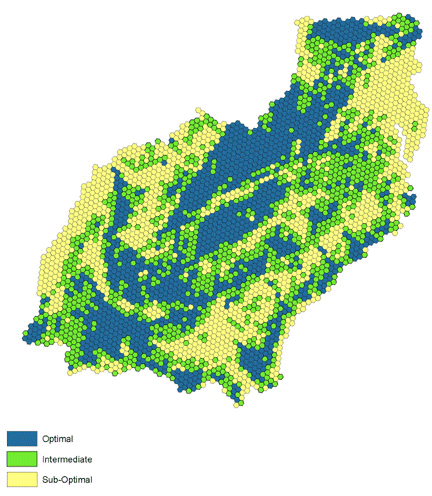
A ranking approach works well in broad-scale environmental analysis (Jones et al. 1997, O’Connell et al. 1998, Wickham et al. 1999). For each habitat variable, we developed an index such that an equal number of hexagons fell into each of three condition classes [optimal (3), intermediate (2), and sub-optimal (1)] and combined all rankings into a single measure. Thus a value of 3 represents sub-optimal stopovers, values 4–6 represent intermediate stopovers, and 7–9 represent optimal stopovers (Fig. 5). We selected hexagons in the “optimal” habitat ranking; all others were classified as non-habitat. At a sub-continental scale, these habitats may provide the core stopovers that ensure long-term successful migration for large populations of birds.
|
Fig. 5. Habitat condition index, based on local forest density, percent agriculture, and road density.
|
Flight algorithms
Most springtime migrants in this study area make landfall along the Gulf and Atlantic coasts. We used every landfall hexagon as the start of a pathway; migrants at the end of a trans-Gulf flight are in a uniquely depleted energetic condition, and may use a remarkable variety of habitats (Moore et al. 1995). Biometric modeling suggests that equal nightly flight distances are the best strategy for minimizing time spent en route, assuming optimal refueling opportunities all along the route (Weber and Houston 1997b). Although we recognize that such idealized flight sequences may not be possible, or even desirable, in the real world (Weber and Houston 1997a), we hypothesize that a landscape capable of sustaining inflexible equal-flight migration can sustain more varied migration strategies as well.
Each modeling iteration represents a different combination of habitat, orientation, and flight distance. We created a movement algorithm within the Arc/Info Geographic Information System (ESRI 2001). Simons et al. (2000) applied flight equations developed by Pennycuick (1989) to data from migrants banded on the Gulf Coast. We averaged their results for forest migrants and modeled flights for migrants in average energetic condition (140–150 km), low energetic condition (90–100 km), and excellent energetic condition (190–200 km). We combine these distances with various orientations: northeast-bound migrants (20–60 degree azimuth), northbound migrants (340–20), and northwest-bound migrants (300–340), yielding a total of nine model iterations.
We use a second modeling approach to assess whether birds with different migration strategies congregate in certain optimal stopovers; birds leaving different areas may arrive at the same stopover after following different pathways with varying flight distances. We started with a narrow orientation window (350–10, 35–55, 305–325 degree azimuth) and then widened the spread by 20 degrees (e.g., 340–20, 25–65, 295–335) for subsequent iterations, to a maximum 80-degree window. Combining each orientation set with varying windows of flight distance (130–150, 120–160, 110–170, 100–180 kilometers), representing birds in various energetic condition, produced maps of overlapping stopover use for birds headed north, northeast, and northwest, plus an overall-use map representing all model iterations.
We ran nine iterations of the fixed distance and orientation models (Table 2). We evaluated each landfall point to determine if the pathway it yielded failed immediately (no additional habitats selected); failed in the first, second, or final third of the pathway (from landfall to northern boundary); successfully completed migration (subsequent flights cross northern boundary); or was inappropriate (e.g., eastern migration from eastern shore points). We found that increases in flight distance (energetic condition) increase success, and northeasterly migrations have significantly more intact pathways for all distances.
|
Table 2. Efficacy of fixed-distance, fixed-orientation migration from landfall points in southeastern North America.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
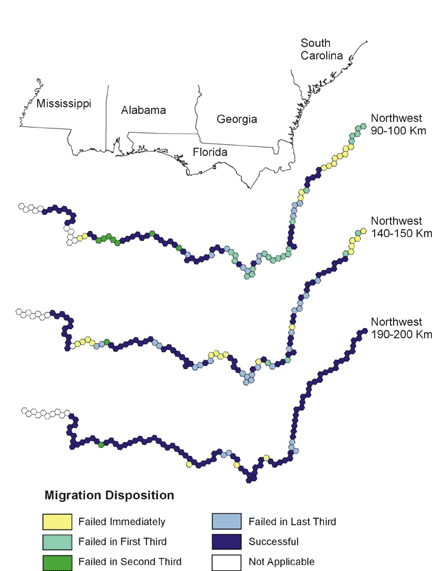
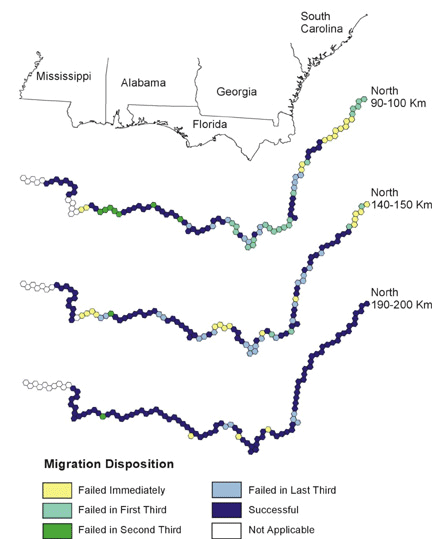
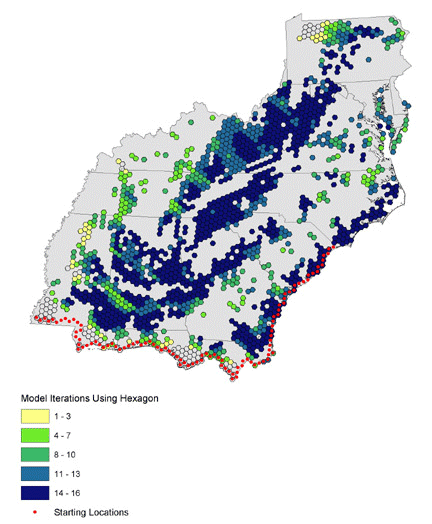
Note the clusters of failed starting points in Figs. 6, 7, and 8, and the reduced size of these clusters as migratory distance increases. Compare these figures with the pattern of optimal habitat (Fig. 9). Sub-optimal areas where large numbers of pathways failed are highlighted with numbered ovals and illustrate the importance of geography in stopover analysis. The spatial distribution of optimal stopovers determines which pathways succeed.
|
Fig. 6. Northwest migration. Colors indicate the modeled ability of birds making landfall to complete migration from that location.
|
|
Fig. 7. North migration. Colors indicate the modeled ability of birds making landfall to complete migration from that location.
|
|
Fig. 8. Northeast migration. Colors indicate the modeled ability of birds making landfall to complete migration from that location.
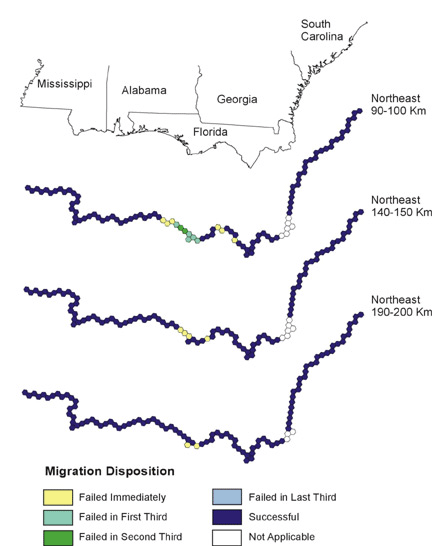
|
|
Fig. 9. Key locations where a lack of optimal habitat results in the failure of modeled migratory pathways.
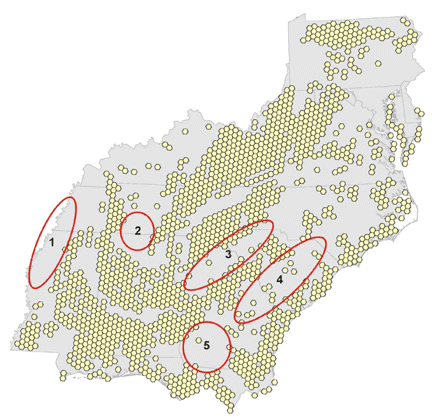
|
Overlapping models
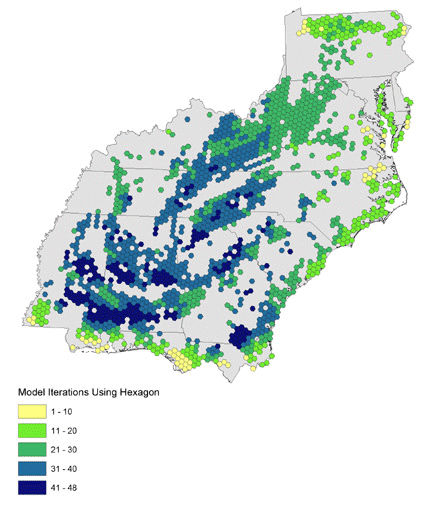
We ran 16 combinations of orientation and nightly flight distance for north-, northeast-, and northwest-bound migrants and present results as the number of models that use each hexagon (Figs. 10, 11, 12). Hexagons where many iterations overlap may represent stopovers that are important to a wide variety of birds in different energetic conditions and flying different routes. North- and northwest-trending birds cluster in the southwest and in identifiable bands through the central portion of the study area. Northeast-trending birds cluster along the northeast–southwest axis of the Appalachians, with a great deal of overlap at many locations across the landscape. We combined all 48 iterations of the collective models to identify locations where birds of all migratory orientations and energetic conditions tend to congregate (Fig. 13). There is a pattern of overlapping bands in the southwest, and it is likely that this trend would continue eastward if not for the lack of habitat at location 5 in Fig. 9. The Appalachians show a strong overlapping pattern, particularly in the south.
|
Fig. 10. Habitat area use modeled for northwest-bound (azimuth 275–355) migration for all distance iterations (100–200 km nightly flights).
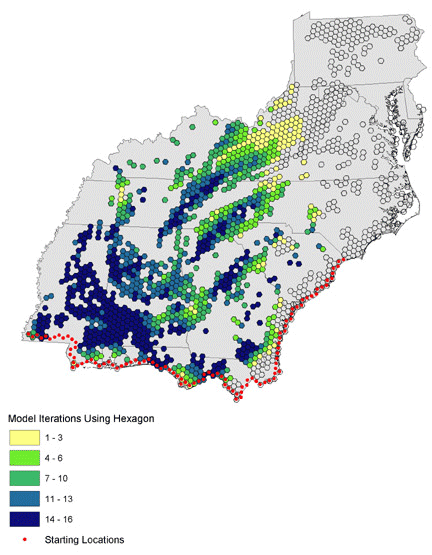
|
|
Fig. 11. Habitat area use modeled for northbound (azimuth 320–40) migration for all distance iterations (100–200 km nightly flights).
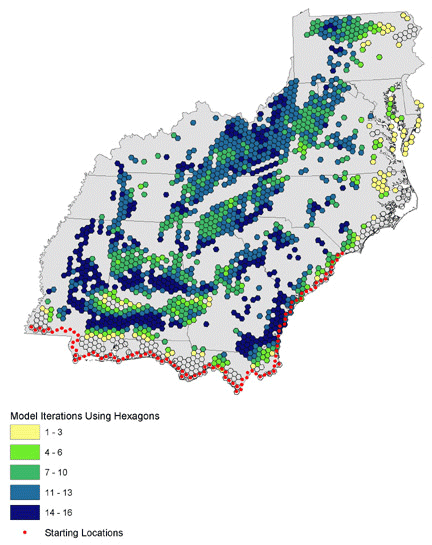
|
|
Fig. 12. Habitat area use modeled for northeast-bound (azimuth 5-85) migration for all distance iterations (100-200 km nightly flights).
|
|
Fig. 13. Modeled habitat area use for all combinations of migration distance and direction.
|
We developed and analyzed speculative models of migration between optimum forest habitats in the eastern United States in an effort to spur geographic migration research and to demonstrate one method of evaluating regional-scale migratory landscapes. Our results suggest that successful pathways can connect remaining optimal stopovers using fixed-distance and fixed-orientation migration, and that geographic location may be as important as habitat condition when evaluating stopover quality. A stopover may have abundant forest habitat and low human disturbance but, if it is not located along a viable migratory corridor, it will not be used. Our models highlight some basic geographic patterns of stopover quality and possible migratory corridors, but these results alone should not be used to assess stopover quality or migration viability at specific locations. Rather, they should be used to examine individual sites within a broad migratory-landscape context. There are many studies of local-scale stopover quality; our goal is to begin to flesh out broad-scale differences between locales. Conservationists and land managers must start to think of habitat in spatial terms, to ensure that stopovers persist all along the migratory pathway at intervals useful to migrants.
Fixed-Distance and Fixed-Orientation MigrationTaking a regional view of migration, landfall points that lead to successful migration corridors may be more important than other landfalls. Although our models are preliminary, it is worth noting that large portions of the Gulf Coast lead to successful pathways for various flight distances (Figs. 6, 7, 8). If our assumptions concerning inherited migration instincts are correct, the landfall point of an individual migrant is determined on initiation of migration from the southern side of the Gulf of Mexico. Migrants may instinctually select a flight direction with no knowledge of what lies at the end of the trans-Gulf flight; thus their entire migration pathway may depend on selecting a flight direction that leads to a successful landfall that, in turn, leads to intact pathways. Assessing the importance of such decisions requires the type of modeling that we illustrate here.
The modern geography of optimal stopovers is composed of multiple “clumps” of high-quality habitat widely dispersed along the migratory route. Birds with higher energetic condition (and longer flights) are better able to cope with this spatial pattern. Longer flights yield a shorter overall migration, which may increase breeding success through acquisition of prime territories; however, dwindling food supply and increased metabolic rate for thermoregulation (Kokko 1999) complicate the benefits of a quick migration. Empirical studies of fat deposits (e.g., Kaiser 1992) and modeling studies based on Pennycuick (1989, 1997) report an average flight distance of 140–150 km for passerine migrants; our findings suggest such a distance is viable for many pathways across the modern habitat landscape.
Past research has noted that the northeast–southwest trending Appalachian Mountains are an important feature of the migration landscape (Berthold 1993), and our models highlight the Appalachians as well. In contrast, the highlighted areas in Figure 9 account for most failed migration pathways. Although it is axiomatic that deforestation affects individual forested stopovers, our results suggest that the broad spatial pattern of deforestation may have an overall direct impact on the efficacy of migratory corridors by removing important steps along the migration route.
Overlapping models of migration direction and durationOur goal in developing the overlapping models was to examine how the current idealized landscape might support large populations of birds with different migratory parameters. Our results suggest that northeast-bound birds are best adapted to the modern landscape; northwest-bound birds remain successful due to fortuitous spacing of habitat “clumps” in the landscape; and northbound birds may be most challenged by the current distribution of stopovers. The patterns seen in Figs. 10, 11, 12, and 13 are created by the overlap of pathways: modeled birds leave from different landfalls, fly along different orientations for different lengths of time, and arrive at the same stopover. These stopovers may provide resiliency to migratory species by accommodating many combinations of orientation and flight distance, preserving the genetic diversity of migrant populations.
We hypothesize that the overlapping pathways seen in our models illustrate a new way of looking at a real phenomenon called broad-front migration: migrating passerines appear to “flow” across regions in broad patterns, showing no preference for specific migration pathways (Hutto 2000, Berthold 1993). Radar images of airborne migrants support this scenario (Tankersley, unpublished data). Although migrants do move in unison, particularly in response to weather patterns, our results suggest they may still be following precise migratory corridors.
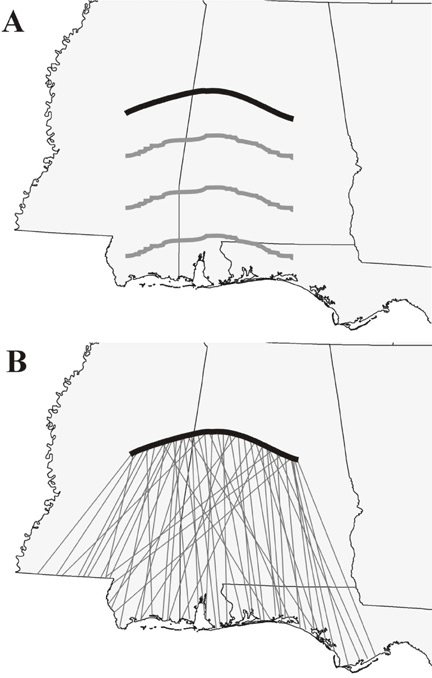
Individual populations moving in parallel may give the appearance of broad-front migration (Fig. 14; also, Berthold 1993), but the difference has profound conservation implications. If broad-front migration implies migrants have no affinity for particular migration corridors, we could reasonably expect to remove an individual stopover from a cluster of stopovers with little effect on any one migrant population. If migrants target discrete, adjacent stopovers, removal may disproportionately impact a specific breeding population. Many studies suggest the heritability of migratory parameters (Berthold 1988, Kerlinger 1995, Hutto 1985), and habitat alteration in this scenario could genetically compress a species toward using fewer migratory corridors. Habitat alteration may then destroy those few corridors and break the connection between wintering and breeding grounds, yielding the geographic patterns of migrant population decline seen in the BBS data (Villard and Maurer 1996). Our results neither prove nor disprove this scenario; rather, they present an alternative mechanism of broad-front migration that merits consideration by migration researchers.
|
Fig. 14. Illustration of broad-front migration (A) and overlapping migration pathways (B).
|
Building more robust models
The models we discuss here simplistically represent complex instincts embodied by migrant birds, and are primitive relative to the journeys made by real birds. This research is a speculative first step that takes a novel look at migration as a process of selecting individual stopovers from the set of stopovers made available by the direction and duration of migratory travel. Our goal was to show that migration geography matters: migrants must be able to find suitable habitats suitably placed all along the migratory path, and regional studies are the only way to evaluate how and where the current landscape provides intact migratory routes. The models we present tentatively illustrate the type of modeling that is needed, and should be seen as a research direction rather than an endpoint. Nonetheless, through our tackling of the broad-scale questions of migration, we are able to recommend several research areas where new models can provide better information:
- Energetic condition and migratory flight distance.—We assume perfect feeding at every location, and static energetic condition. Real habitats harbor different feeding opportunities, yielding different refueling rates, and future models should account for nightly changes in flight distance capability. Differences in refueling rates are both random (habitats at any location may be optimal or poor) and systematic (food availability in spring decreases monotonically further northward), and human development degrades habitats in predictable patterns (Alerstam and Hedenstrom 1998). These conditions can be modeled.
- Complex navigation.—Birds probably supplement their inherited migratory orientation with visual cues and knowledge of specific locations. Leading lines in the landscape (such as river corridors, mountain ranges, and coastlines) may augment innate directional cues as birds start out on a compass bearing and
then follow visible landmarks (Moore and Aborn 2000). Real birds may also use a disjunct mental map that joins disparate habitats with a known flight distance to create a less complex, yet complete, mental image of the landscape (Berthold 1996, Baker 1984). Different model scenarios could account for specific physiography and explore how mountain ranges, large water bodies, or sprawling metropolitan areas might influence constant orientation pathways.
- Weather.—Prevailing winds affect the overall location of migratory pathways, and storms may temporarily alter individual paths (Moore et al. 1993, Moore and Simons 1992, Richardson 1991). Migrants may compensate for wind drift both on the wing and during morning flights, ultimately yielding constant orientation pathways (Berthold 1996, see also Richardson 1991). It is unknown if weather events can override innate cues for habitat, direction, and duration. We need more research on the broad-scale implications of weather patterns during migration, and how those patterns may affect the locations of migratory pathways.
- Model scale and landscape representation.—Broad habitat conditions may inform migrants when selecting one stopover from a collection of nearby stopovers. Our models use hexagons similar in scale to other landscape ecology studies of breeding birds. Given current computing power and more robust statistical techniques, such as Mahalanobis distance modeling, scientists should be able to refine the habitat picture to capture meso-scale differences in habitats
yet retain an overall regional perspective. This requires field research at many locations and modeling research to bring results from different areas together into a coherent migration geography.
- Map projections.—Birds may follow great circle (orthodrome) routes that conform to the shape of the earth (Alerstam and Hedenstrom 1998). An orthographic projection captures these routes and could help us explore orthodromes and migration along magnetic declination lines. The spatial coordinate system of orthographic projections, however, is computationally intensive and requires cutting-edge computer technology. As Gudmundsson and Alerstam (1998) note, the use of different map projections may be a powerful tool for examining assumptions about migratory orientation.
CONCLUSION Our speculative models embody some of the predominant theories in migration literature, and suggest a complex landscape governed by inherited migration strategies and multi-scale characteristics of available habitats. We hope that our approach and preliminary results will encourage further broad-scale migration studies, both of forest migrants and other migratory bird species. The geography of remaining habitats is important, and effective conservation must consider how each habitat fits into the overall landscape of migration pathways. Disjunct studies of individual stopovers, with no effort to integrate findings across the landscape, are insufficient to guarantee successful migration for neotropical birds. Future research must strive to combine on-site study with broad-scale mapping to create a unified migration landscape, then study how birds move across that landscape and identify locales where conservation or restoration is needed. Perhaps new technologies, such as satellite tracking (Nowak and Berthold 1991) and isotope analysis (Marra et al. 1998), will provide hard evidence of pathways connecting specific stopovers. Until then, we must continue with focused projects that use spatial models embodying information from many stopovers to test assumptions and refine migration theory.
RESPONSES TO THIS ARTICLE Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a comment, follow this link. To read comments already accepted, follow this link.
Acknowledgments: This research was funded in part by the Environmental Protection Agency through an interagency agreement with Tennessee Valley Authority. Both Nikon Corporation and Bausch and Lomb Corporation provided reduced-cost optics for field study. The authors thank Drs. Carol Harden, Sally Horn, Daniel Simberloff, and anonymous reviewers for suggestions that improved this manuscript.
APPENDIX 1 Explanation of sliding window algorithms.
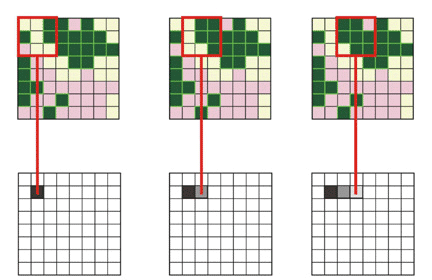
The National Pattern Database (Riitters et al. 2000a) provides maps of local forest density based on computational analysis of the National Landcover Dataset (Loveland and Shaw 1996). Riitters used a cell-by-cell computation algorithm within a sliding window to evaluate local forest density, with a window size of 65 ha, encompassing 729 30-m pixels (picture elements or mapping units). This algorithm evaluates the landscape in the analysis window, computes the forest landcover measure, and tabulates the result for each point, yielding a continuous surface map of the region.
APPENDIX 2 Illustration of sliding window algorithm.
LITERATURE CITED Able, K. P. 1972. Fall migration in coastal Louisiana and the evolution of migration patterns in the Gulf region. Wilson Bulletin 84:231–242.
Alerstam, T., and A. Hedenstrom. 1998. The development of bird migration theory. Journal of Avian Biology 29:343–369.
Askins, R. A., J. F. Lynch, and R. Greenberg. 1990. Population declines in migratory birds in eastern North America. Current Ornithology 7:1–57.
Baker, R. R. 1984. Bird navigation: the solution of a mystery? Holmes and Meier, New York, New York, USA.
Berthold, P. 1988. The control of migration in European warblers. Acta XIX Congressus Internationalis Ornithologici 19:215–249.
Berthold, P. 1990. Genetics of migration. Pages 269–280 in E. Gwinner (editor). Bird migration: the physiology and ecophysiology. Springer, Berlin, Germany.
Berthold, P. 1991. Orientation in birds. Birkhäuser, Boston, Massachussetts, USA.
Berthold, P. 1993. Bird migration: a general survey. Oxford University Press, New York, New York, USA.
Berthold, P. 1996. Control of Bird Migration. Chapman and Hall, London, UK.
Berthold, P., A. J. Helbig, G. Mohr, and U. Querner. 1992. Rapid microevolution of migratory behavior in a wild bird species. Nature 360:668–669.
Bingman, V. P., K. P. Able, and P. Kerlinger. 1982. Wind drift, compensation, and the use of landmarks by nocturnal bird migrants. Animal Behavior 30:49–53.
Brawn, J. D., and S. K. Robinson. 1996. Source–sink population dynamics may complicate the interpretation of long-term census data. Ecology 77(1):3–12.
Bruderer, B., and L. Jenni. 1990. Migration across the Alps. Pages 60–77 in E. Gwinner, editor. Bird migration: the physiology and ecophysiology. Springer-Verlag, Berlin, Germany.
Cochran, W. W., and C. G. Kjos. 1985. Wind drift and migration of thrushes: a telemetry study. Illinois Natural History Survey Bulletin 33:297–330.
Cochran, W. W., G. G. Montgomery, and R .R. Graber. 1967. Migratory flights of Hylocichla thrushes in spring: a radiotelemetry study. Living Bird 6:213–225.
Dunn, J., and K. Garrett. 1997. A field guide to warblers of North America. Houghton Mifflin, New York, New York, USA.
ESRI. 1997. United States street database. Earth Systems Research Institute, Redlands, California, USA.
ESRI. 2001. Arc/Info geographic information system. Earth Systems Research Institute, Redlands, California, USA.
Finch, D. M. 1991. Population ecology, habitat requirements, and conservation of neotropical migratory birds. U. S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station, General Technical Report RM–205, Fort Collins, Colorado, USA.
Flather, C. H., and J. R. Sauer. 1996. Using landscape ecology to test hypotheses about large-scale abundance patterns in migratory birds. Ecology 77(1):28–35.
Freemark, K., and B. Collins. 1992. Landscape ecology of birds breeding in temperate forest fragments. Pages 443–454 in J. M. Hagan, III and D. W. Johnston, editors. Ecology and conservation of neotropical migrant landbirds. Smithsonian Institution Press, Washington, D. C., USA.
Gudmundsson, G. A., and T. Alerstam. 1998. Optimal map projections for analysing long-distance migration routes. Journal of Avian Biology 29:597–605.
Hagstrum, J. 2000. Infrasound and the avian navigational map. Journal of Experimental Biology 203:1103–1111.
Hall, G. A. 1981. Fall migration patterns of wood warblers in the southern Appalachians. Journal of Field Ornithology 52(1):43–49.
Hedenstrom, A., and T. Alerstam. 1998. How fast can birds migrate? Journal of Avian Biology 29: 424–432.
Hunter, W. C., D. N. Pashley, and R. E. F. Escano. 1993. Neotropical migratory landbird species and their habitats of special concern within the Southeast region. Pages 159–171 in D. M. Finch and P. W. Stangel, editors. Status and management of neotropical migratory birds. U. S. Department of Agriculture Forest Service Technical Report RM–229. Fort Collins, Colorado, USA.
Hutto, R. L. 1985. Habitat selection by nonbreeding, migratory land birds. Pages 455–476 in M. Cody, editor. Habitat election in birds. Academic Press, New York, New York, USA.
Hutto, R. L. 2000. On the importance of en route periods to the conservation of migratory landbirds. Studies in Avian Biology 20:109–114.
Jones, K. B., K. H. Riitters, J. D. Wickham, R. D. Tankersley, R. V. O’Neill, D. J. Chaloud, E. R. Smith, and A. C. Neale. 1997. An ecological assessment of the United States mid-Atlantic region: a landscape atlas. EPA/600/r-97/130. Office of Research and Development, Washington, D. C., USA.
Kaiser, A. 1992. Fat deposition and theoretical flight range of small autumn migrants in southern Germany. Bird Study 39:96–110.
Keast, A., and E. S. Morton, editors. 1980. Migrant birds in the neotropics: ecology, behavior, distribution, and conservation. Smithsonian Institution Press, Washington, D. C., USA.
Kelly, J. F., R. Smith, D. M. Finch, F. R. Moore, and W. Yong. 1999. Influence of summer biogeography on wood warbler stopover abundance. Condor 101:76–85.
Kerlinger, P. 1995. How birds migrate. Stackpole Books, Mechanicsburg, Pennsylvania, USA.
Kerlinger, P., and F. R. Moore. 1989. Atmospheric structure and avian migration. Current Ornithology 6:109–142.
Kokko, H. 1999. Competition for early arrival in migratory birds. Journal of Animal Ecology 68:940–950.
Loveland, T. R., and D. M Shaw. 1996. Multiresolution land characterization: building collaborative partnerships. In J. M. Scott, T.Tear, and F. Davis, editors. Gap analysis: a landscape approach to biodiversity planning. Proceedings of the ASPRS/GAP Symposium, Charlotte, North Carolina, USA. National Biological Survey, Moscow, Idaho, USA. pp. 83–89.
Marra, P. P., K. A. Hobson, and R. T. Holmes. 1998. Linking winter and summer events in a migratory bird by using stable-carbon isotopes. Science 282:1884–1886.
Martin, G. R. 1990. The visual problems of nocturnal migration. Pages 187–197 in E. Gwinner, editor. Bird migration: the physiology and ecophysiology. Springer, Berlin, Germany.
Moore, F. R. 1984. Age-dependent variability in the orientation of migratory savannah sparrows (Passerculus sandwichensis). Auk 101:875–880.
Moore, F. R., and D. A. Aborn. 2000. Mechanisms of en route habitat selection: how do migrants make habitat decisions during stopover? Studies in Avian Biology 20:34–42.
Moore, F. R., and P. Kerlinger. 1987. Stopover and fat deposition by North American wood-warblers (Parulinae) following spring migration over the Gulf of Mexico. Oecologia 74:47–54.
Moore, F. R., and T. R. Simons. 1992. Habitat suitability and stopover ecology of neotropical landbird migrants. Pages 345–355 in J. M. Hagan III and D. W. Johnston, editors. Ecology and conservation of neotropical migrant landbirds. Smithsonian Institution Press, Washington, D. C., USA.
Moore, F. R., S. A. Gauthreaux, Jr., P. Kerlinger, and T. R. Simons. 1993. Stopover habitat: management implications and guidelines. Pages 58–69 in D. M. Finch and P. W. Stangel, editors. Status and management of neotropical migratory birds. U. S. Department of Agriculture Forest Service Technical Report RM–229. Fort Collins, Colorado, USA.
Moore, F. R., S. A. Gauthreaux, Jr., P. Kerlinger, and T. R. Simons. 1995. Habitat requirements during migration. Pages 121–144 in T. E. Martin and D. M. Finch, editors. Ecology and management of neotropical migratory birds. Oxford University Press, New York, New York, USA.
Nicholson, C. P. 1997. Atlas of the breeding birds of Tennessee. University of Tennessee Press, Knoxville, Tennessee, USA.
Nisbet, I. C. T. 1969. Returns of transients: results of an inquiry. EBBA News 32:269–274.
Nowak, E., and P. Berthold. 1991. Satellite-tracking: a new method in orientation research. Pages 86–104 in P. Berthold, editor. Orientation in birds. Birkhäuser, Boston, Massachussetts, USA.
O’Connell, T. J., L. E. Jackson, and R. P. Brooks. 1998. A bird community index of biotic integrity for the mid-Atlantic Highlands. Environmental Monitoring and Assessment 51:145–156.
Papi, F. 1991. Olfactory navigation. Pages 52–85 in P. Berthold, editor. Orientation in birds. Birkhäuser, Boston, Massachussetts, USA.
Pennycuick, C. J. 1989. Bird flight performance: a practical calculation manual. Oxford University Press, New York, New York, USA.
Pennycuick, C. J. 1997. Actual and optimum flight speeds: field data reassessed. Journal of Experimental Biology 200:2355–2361.
Petit, D. R. 2000. Habitat use by landbirds along nearctic–neotropical migration routes: implications for conservation of stopover habitats. Studies in Avian Biology 20:15–33.
Rappole, J. H. 1995. The ecology of migrant birds: a neotropical perspective. Smithsonian Institution Press, Washington, D. C., USA.
Richardson, W. J. 1991. Wind and orientation of migrating birds: a review. Pages 226–249 in P. Berthold, editor. Orientation in birds. Birkhäuser, Boston, Massachussetts, USA.
Riitters, K. H., J. D. Wickham, J. E. Vogelmann, and K. B. Jones. 2000. National landcover pattern data. Ecology 81:604–607.
Robbins, C. S., S. Droege, and J. R. Sauer. 1989. Monitoring bird populations with Breeding Bird Survey and atlas data. Annales Zoologica Fennici 26:297–304.
Sandberg, R., and F. Moore. 1996. Migratory orientation of red-eyed vireos, Vireo olivaceus, in relation to energetic condition and ecological context. Behavioral Ecology and Sociobiology 39:1–10.
Sandberg, R., J. Pettersson, and T. Alerstam. 1988. Why do migrating robins, Erithacus rubecula, captured at two nearby stop-over sites orient differently? Animal Behavior 36:865–876.
Sandberg, R., J. Petterson, and K. Persson. 1991. Migratory orientation of free-flying robins Erithacus rubecula and pied flycatchers Ficedula hypoleuca: release experiments. Ornis Scandinavica 22:1–11.
Schmidt-Koenig, K., J. U. Ganzhorn, and R. Ranvaud. 1991. The sun compass. Pages 1–15 in P. Berthold, editor. Orientation in birds. Birkhäuser, Boston, Massachussetts, USA.
Simons, T. R., S. M. Pearson, and F. R. Moore. 2000. Application of spatial models to the stopover ecology of trans-Gulf migrants. Studies in Avian Biology 20:15–33.
Tankersley, R., Jr. 2002. Geography of stopover habitats and migratory pathways for neotropical migratory forest birds in the eastern United States. Dissertation, University of Tennessee, Knoxville, Tennessee, USA.
Villard, M., and B. A. Maurer. 1996. Geostatistics as a tool for examining hypothesized declines in migratory songbirds. Ecology 77(1):59–68.
Weber, T. P., and A. I. Houston. 1997a. A general model for time-minimising avian migration. Journal of Theoretical Biology 185:447–458.
Weber, T. P., and A. I. Houston. 1997b. Flight costs, flight range and the stopover ecology of migrating birds. Journal of Animal Ecology 66:297–306.
Wickham, J. D., K. B. Jones, K. H. Riitters, R. V. O’Neill, R. D. Tankersley, E. R. Smith, A. C. Neale, and D. J. Chaloud. 1999. An integrated environmental assessment of the U. S. mid-Atlantic region. Environmental Management 24:553–560.
Wiltschko, W., and R. Wiltschko. 1988. Magnetic orientation in birds. Current Ornithology 5:67–121.
Wiltschko, W., and R. Wiltschko. 1991. Magnetic orientation and celestial cues in migratory orientation. Pages 16–37 in P. Berthold, editor. Orientation in birds. Birkhäuser, Boston, Massachussetts, USA.
Winker, K., D. W. Warner, and A. R. Weisbrod. 1991. Unprecedented stopover site fidelity in a Tennessee warbler. Wilson Bulletin 103:514–516.
Winker, K., D. W. Warner, and A. R. Weisbrod. 1992. Daily mass gains among woodland migrants at an inland stopover site. Auk 109(4):853–862.
Woodrey, M. S. 2000. Age-dependent aspects of stopover biology of passerine migrants. Studies in Avian Biology 20:43–52.
Woodrey, M. S., and F. R. Moore. 1997. Age-related differences in the stopover of fall landbird migrants on the coast of Alabama. Auk 114:695–707.
Yong, W., D. M. Finch, F. R. Moore, and J. F. Kelly. 1998. Stopover ecology and habitat use of migratory Wilson’s warblers. Auk 115(4):829–842.
Address of Correspondent:
Roger Tankersley, Jr.
Tennessee Valley Authority,
P. O. Box 1649,
Norris, Tennessee 37828 USA
Phone: (865)632-1427
Fax: (865)632-1493
rdtankersley@tva.gov