|
|
|
Copyright ©1998 by The Resilience Alliance*
Host, G. and J. Pastor. 1998. Modeling forest succession among ecological land units in northern Minnesota. Conservation Ecology [online] 2(2): 15. Available from the Internet. URL: http://www.consecol.org/vol2/iss2/art15/
A version of this article in which text, figures, tables, and appendices are separate files may be found by following this link.
Research, part of Special Feature on Sustainability and Resilience in Boreal Regions Modeling Forest Succession among Ecological Land Units in Northern Minnesota George Host and John Pastor
Natural Resources Research Institute, University of Minnesota - Duluth
- Abstract
- Introduction
- Methods
- Results
- Discussion
- Responses to This Article
- Acknowledgments
- Literature Cited
Field and modeling studies were used to quantify potential successional pathways among fine-scale ecological classification units within two geomorphic regions of north-central Minnesota. Soil and overstory data were collected on plots stratified across low-relief ground moraines and undulating sand dunes. Each geomorphic feature was sampled across gradients of topography or soil texture. Overstory conditions were sampled using five variable-radius point samples per plot; soil samples were analyzed for carbon and nitrogen content. Climatic, forest composition, and soil data were used to parameterize the sample plots for use with LINKAGES, a forest growth model that simulates changes in composition and soil characteristics over time. Forest composition and soil properties varied within and among geomorphic features. LINKAGES simulations were using "bare ground" and the current overstory as starting conditions. Northern hardwoods or pines dominated the late-successional communities of morainal and dune landforms, respectively. The morainal landforms were dominated by yellow birch and sugar maple; yellow birch reached its maximum abundance in intermediate landscape positions. On the dune sites, pine was most abundant in drier landscape positions, with white spruce increasing in abundance with increasing soil moisture and N content. The differences in measured soil properties and predicted late-successional composition indicate that ecological land units incorporate some of the key variables that govern forest composition and structure. They further show the value of ecological classification and modeling for developing forest management strategies that incorporate the spatial and temporal dynamics of forest ecosystems.
KEY WORDS: climatology, dunes, ecological classification systems (ECS), forest growth model, forest management, forest, succession, geomorphology, LINKAGES, moraines, northern Minnesota, overstory composition, overstory composition, soil properties.
Ecological Classification Systems (ECS) provide a means to quantify variation in forest composition, productivity, and fundamental ecological processes across regional landscapes. Specifically, the map units defined in an ECS delimit stable abiotic components of ecosystems that, when used in conjunction with existing biota, represent landscape ecosystems that recur in characteristic physiographic positions, are relatively homogeneous in terms of structure and function, and exhibit similar responses to management activities or natural disturbances (Barnes et al. 1982). As such, ECS provides a means to understand how different parts of the landscape respond to management, and thereby improve both strategic and tactical planning for managing natural resources in a sustainable and environmentally sound manner.
A fundamental step in the development and use of an ECS is the interpretation of ecological land units in terms of specific management objectives. These objectives include assessments of forest succession (Host et al. 1987), response to silvicultural treatments, productivity (Host et al. 1988), species diversity (Host and Pregitzer 1991), game and nongame habitat (Johnson et al. 1991), operability, and others. Each of these objectives requires a description or characterization of an ecosystem, in terms of its composition and structure (e.g., species composition, physiognomy, wildlife habitat), as well as ecosystem functional processes (e.g., nutrient cycling, productivity, or succession). The compositional and structural attributes of an ecosystem are often described from data used in ECS development and supplemented by data collected for ECS validation (Host et al. 1993). To characterize ecosystem processes, however, often requires explicit field or modeling studies apart from ECS development.
The objective of this study was to use field and modeling studies to quantify potential successional pathways among fine-scale ecological classification units within two geomorphic regions in the Chippewa National Forest, north-central Minnesota. Modeling was based on LINKAGES, a gap model that simulates species replacement patterns as a function of species life history attributes interacting with soil nutrient dynamics (Pastor and Post 1986). The simulation runs were parameterized based on soil and overstory data collected in the field. Simulations were run from bare-ground and current conditions to assess degree of convergence or divergence of forest composition among ecosystems. Results were used to suggest refinements to current ecological land classification practices, as well as to address the use of simulation modeling tools in forest management.
Field sample plots were stratified across the landscape, based on the existing ecological classification system for the Chippewa National Forest in north-central Minnesota (Shadis et al. 1995). Plots were stratified across two major geomorphic units on the Forest; these units are referred to in ECS terminology as LandType Associations or LTAs. The Guthrie Till Plain (LTA E) is a low-relief ground moraine consisting of medium-textured till. The soils are predominantly sandy loams underlain by clay loams. The Bena Dunes (LTA O) consist of sand dunes with varying depths to the regional water table. Differences in slope position and depth to water table allow the identification of finer scaled classification units, known as Ecological Landtype Phase (ELTPs), or simply, Phases. Within each LTA, we sampled three dominant Phases that span the upland soil moisture gradient, from very dry sites in high landscape positions to sites that were somewhat poorly drained (LTA E) or characterized by high seasonal water table (LTA O; Table 1).
Table 1. Ecological Landtype Phases (ELTP) and soil moisture characteristics among Landtype Associations.
| ||||||||||||||||||||
Field and laboratory methods
Five sample plots were located at random within each ELTP, resulting in a total number of 30 sample plots (5 plots x 3 ELTPs x 2 LTAs). Within each plot, five variable-radius point samples (10 BAF) were taken, and the species and diameter at breast height (dbh) of each tally tree were recorded. Heights and ages were recorded on three trees at each point.
Three soil cores were taken at each point to determine soil carbon and nitrogen contents. Four additional cores were collected at intermediate distances between sample points to more fully characterize variation within the stand, resulting in 19 samples per plot. Soils were collected to a depth of 50 cm using a standard soil probe. Samples were mixed thoroughly and air-dried. Each of the 570 soil samples was analyzed for percentage of carbon and nitrogen (by mass) using a CHN analyzer. Bulk density was estimated as a function of soil carbon (Grigal et al. 1989), and was used to convert percentages to C and N content.
Data analysisForest composition was assessed using both univariate and multivariate statistics. Univariate statistics included LTA and ELTP-level summaries of basal area and frequency of occurrence. Patterns of community composition were assessed using Detrended Correspondence Analysis (DCA), a multivariate technique that ordinates sample stands based on similarities in forest composition (Hill 1979).
Simulation methodsPlot-level overstory basal area and soils data were used to parameterize the LINKAGES model (Pastor and Post 1986), which simulates changes in soils and forest composition over time. Model runs were conducted from bare-ground conditions, and by using the current mean stand conditions as starting points. In the latter case, the model was run from bare ground up to the present forest conditions to generate a realistic forest floor layer. These forest floor estimates were then used as inputs to the present forest condition (Pastor and Post 1986).
Climatic inputs for the model consist of mean monthly precipitation and temperature, along with their standard deviations. These weather inputs for the study sites were derived from interpolated estimates of 30-yr average temperature and precipitation data (Host et al. 1995). Regional averages for these variables were derived from a recently developed climatic classification of Minnesota, based on these interpolated data (ZedX 1995).
The standard deviations associated with climatic data were used to simulate the natural variation in weather for each month. As a result, simulated weather and, thus, predicted growth, differs in each year of the model run. To obtain estimates of the average conditions, a series of replicate runs was conducted, and average results were calculated. Results from each plot were an average of 10 replicate simulations.
Several phases on the Bena Dunes are characterized by depth to water table. To simulate differences in moisture availability as a function of water table, the absolute values of precipitation were decreased and standard deviations were increased during selected months on Phases c and f; this allowed a greater percentage of the growing season to be under drought stress in these phases, with concomitant effects on species growth and mortality.
Model outputs were evaluated with respect to the absolute and relative basal areas of individual tree species, the structural diversity of the forest canopy, and the long-term nitrogen dynamics of the soil. The simulations predict succession in its traditional sense (i.e., compositional changes over time), but also some of the causal factors (nitrogen dynamics) and biotic outcomes (avian habitat diversity).
Forest composition varied at both the LTA and ELTP scale. LTA O was dominated by red and white pines (Pinus resinosa and P. strobus, respectively), trembling aspen (Populus tremuloides), or black ash (Fraxinus nigra), whereas LTA E was dominated by primarily by northern hardwoods: sugar maple (Acer saccharum), basswood (Tilia americana), paper birch (Betula papyrifera), and trembling aspen (Table 2). Within each LTA, there were gradients of species composition related to the moisture and fertility status of the site. Phase c, the driest phase of the Bena Dunes, was dominated primarily by red pine (24 m2/ha basal area) and secondarily by white pine (4 m2/ha). Phase f, a wet-mesic phase of the Bena Dune field, was characterized by saturated lower soil horizons during some part of the growing season. This ELTP had a greater species evenness, with stronger components of red maple (Acer rubrum), trembling aspen, and paper birch as codominants (Table 2). Phase e, the wettest phase of this catena, had two characteristic cover types: black ash or a mix of trembling aspen and balsam fir (Abies balsamea).
Table 2. Basal areas (m2/ha) and frequencies of dominant tree species by Ecological Landtype Phase.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
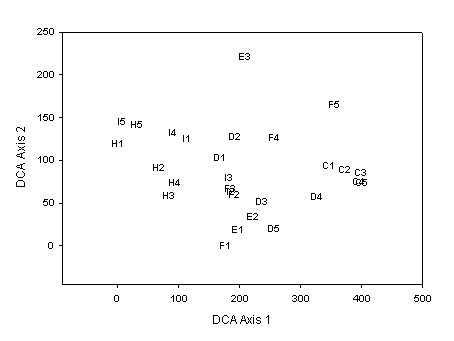
The gradients in species composition just described were also shown in a detrended correspondence analysis (DCA) based on species basal area. On the first DCA Axis, species characteristic of dry sites (i.e., red and white pine) received high scores, whereas northern hardwood species received low scores. Thus, plots characterized by pine received high first-axis scores; among these were plots in Phase c, which were tightly clustered (very similar in species composition), as well as a few plots in Phases f and d (Fig. 1). Phases h and i, which form a catena on upland sites of the Guthrie Till Plain, dominated the low end of Axis 1. Plots of Phase h showed a high dominance by sugar maple (12 m2/ha), whereas Phase i plots were more diverse, with paper birch, sugar maple, aspen, and basswood being codominant. This increased diversity is reflected in the somewhat higher scores for Phase i on the first or second axes. Phase d, which has a significant component of pine, was compositionally similar to some of the plots on Phases e and f of LTA O. Finally, the two plots of Phase e that were characterized by black ash were clearly outliers (as evident in an initial ordination), and were excluded from this DCA. Phase e has since been divided into two Phases, q and l, characterized by maple/birch or black ash, respectively.
Fig. 1. Detrended correspondence analysis (DCA) ordination of 30 field plots, based on overstory basal area. Value labels indicate ELTP and plot number.
|
Soils
Although within-plot variability was high, there were distinct trends in soil C among ELTPs. In LTA O, soil carbon increased with soil moisture content. Soil carbon in Phase c, the driest phase sampled in LTA O, averaged 67 Mg/ha, compared with 96 Mg/ha in the more mesic Phase f (Table 3). Phase e, which occurs under poorly drained conditions, had the highest soil carbon, averaging 148 Mg/ha. However, these high values for Phase e are due, in part, to exceptionally high values of carbon in the two sampled plots dominated by black ash; this also resulted in a much larger variation within this phase. In LTA E, soil carbon was highest in Phase i (112 Mg/ha), which occupies an intermediate landscape position. Carbon averaged 105 Mg/ha in Phase h and 98 Mg/ha in Phase d.
Table 3. Mean and standard deviation of soil carbon and nitrogen among Ecological Landtype Phases.
|
Variability in soil nitrogen was quite high in all phases, and no strong trends were observed; mean nitrogen levels ranged between 5.4 and 6.7 Mg/ha (Table 3). Like carbon, the quantity of nitrogen present in the soil is closely related to the interactions of forest composition and landform. In terms of predicting nutrient availability, and specifically in terms of modeling forest growth, the release of nutrients is more closely related to the C:N ratio than to the absolute values of these variables alone, and C:N showed strong differences among phases.
Simulations
Simulations from bare-ground conditions
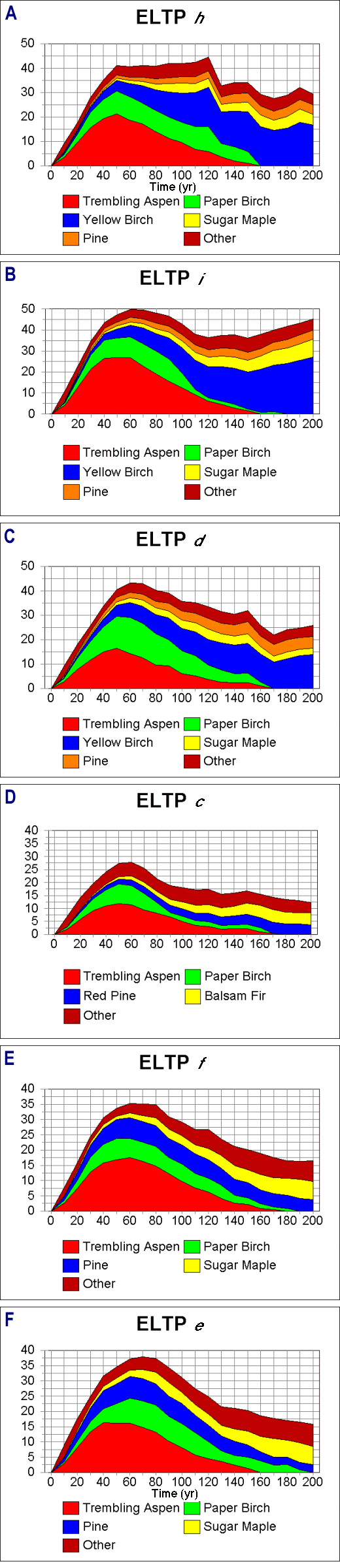
The "bare-ground" runs indicate the general productivity and typical early-successional characteristics of a site as a function of soil texture, C, and N under a particular climatic regime. Productivity levels were generally higher on LTA E than on LTA O, although there was also significant within-LTA variation. All phases showed a significant initial pulse of trembling aspen and, to some degree, paper birch. Species succeeding aspen varied by LTA, with yellow birch and sugar maple being important on LTA E, and red, white, or jack pine becoming more dominant on the drier, sandier LTA O (Fig. 2).
Fig. 2. Simulated forest successional patterns, given
bare-ground starting conditions for six Ecological Landtype Phases on
the Chippewa National Forest, north-central Minnesota. The scale
numbers on the y - axes respresent basal area in
m2/ha.
|
LTA E
On LTA E, all phases were dominated by Prunus pensylvanica in the first 10 yr; this species disappeared by age 30 yr. Phases h and i showed peaks in aspen and birch basal area at age 50 yr, followed by a slow decline in these species as the sites succeeded to more shade-tolerant species (Fig. 2). In the undisturbed conditions of this model run, yellow birch was the major mid-to-late successional species, making up about half of the basal area after age 120 yr. Sugar maple was an important secondary dominant on both h and i, with about twice as much basal area on Phase i (8.6 vs. 4.5 m2/ha) at the end of the run. Total basal area began to level off at age 50 yr in Phase h, followed by a sharp decline at age 120 yr as paper birch began to drop out of the stand. From age 120 to 200 yr, mean total basal area averaged about 30 m2/ha. Phase i showed a similar pattern, although following birch decline, there was a net increase in basal area as yellow birch increased in dominance. Thus, Phase i was characterized by more rapid mortality in early successional species, followed by an accrual in biomass. Pine was a significant and persistent codominant in both of these phases.
Aspen was much less important on Phase d, with paper birch accounting for about half of the initial increase in biomass. Yellow birch was a dominant species toward the end of the run, although it had lower basal area compared with the drier phases. Sugar maple was less important and pine was more important in comparison with Phases h and i. Phase d also showed a stronger decline after peak basal area was reached at year 70.
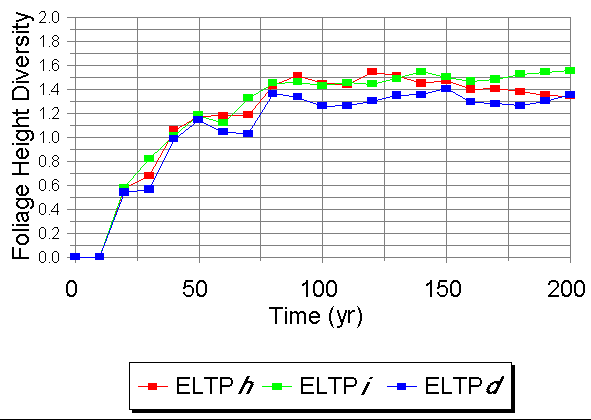
Foliage Height Diversity (FHD), an index of structural diversity that has been shown to be correlated with habitat suitability for a number of bird species (MacArthur 1965), was highest in Phases h and i, with Phase d being consistently lower over the simulation (Fig. 3). Phase i generally had a higher FHD than did Phase h.
Fig. 3. Temporal patterns of Foliage Height Diversity on Ecological Landtype Phases within the Guthrie Till Plain (LTA E) under bare-ground starting conditions.
|
LTA O
Under the relatively dry climate used in this set of simulations, basal areas on Phase c were substantially lower than on Phasesf and e (Fig. 2). In particular, the initial pulse of aspen reached 12 m2/ha on Phase c, compared with 16 and 17 m2/ha on Phases f and e, respectively. In addition, there were compositional differences among the phases, with late succession in Phase c being dominated by red pine and balsam fir; Phase f dominated by maple, fir, and pine; and Phase e including black ash as a component.
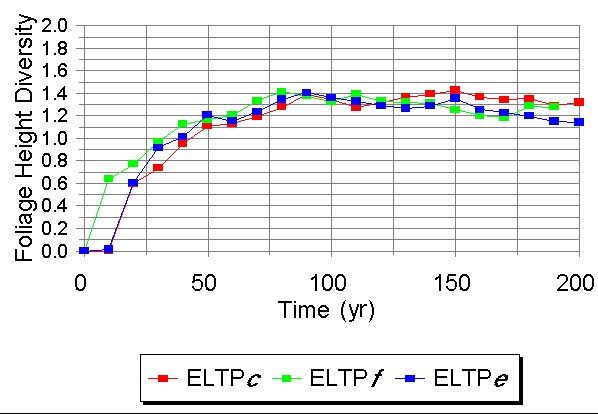
Foliage Height Diversity was highest in Phase f during the biomass acquisition phase of growth (Fig. 4). After biomass reached a maximum, however, Phase c maintained a slightly higher diversity in vertical structure.
Fig. 4. Temporal patterns of Foliage Height Diversity on Ecological Landtype Phases within the Bena Dunes (LTA O) under bare-ground starting conditions.
|
Simulations using current overstory conditions
The current overstory simulations project future compositional patterns, given the current stand conditions in the replicate plots within each phase. As with the bare-ground simulations, there were major differences among LTAs and more subtle differences among ELTPs. The morainal landform showed a decline in mid-successional species such as aspen and birch, and an increase in shade-tolerant species, such as yellow birch, white spruce, and sugar maple. A range of responses was observed in the Bena Dunes, particularly with the pines, although white spruce again was one of the dominant species in late succession.
On the Guthrie Till Plain, many of the plots had a strong initial component of paper birch (Table 2); trembling aspen, a major species in the early-successional runs, was not a strong dominant in the field plots. From the mean starting conditions of the model, paper birch reached a peak basal area about 40 yr into the run and then began a gradual decline, disappearing at age 160 yr (Fig. 5). It is important to note that this is not a single cohort of trees, but the progression of growth within several diameter classes, possibly including species that may have seeded in early in the run. Red and white pine basal areas were greatest on Phase d and decreased through Phases i and h. In all cases, pine declined at a relatively slow rate over the course of the run, probably as a result of the increased shading and changes in forest floor litter quality. The dominant late-successional species was yellow birch, which constituted 46%, 52%, and 39% of total basal area in Phases h, i, and d, respectively. Sugar maple maintained a relatively consistent basal area throughout the run, averaging 5.5 m2/ha in phases h and i, and 4.4 m2/ha in Phase d. White spruce (Picea glauca), a minor component of the initial stand, slowly increased in dominance throughout the run, and constituted between 6.4 and 7.6 m2/ha at the end of the run.
Fig. 5. Simulated forest successional patterns, given current mean overstory conditions for six Ecological Landtype Phases on the Chippewa National Forest.
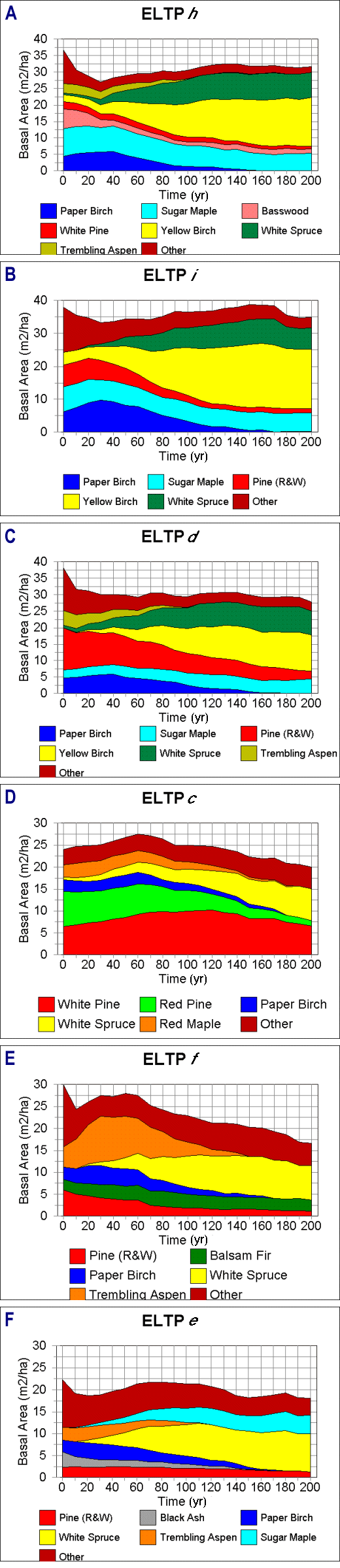
|
On the Bena Dunes, pine exhibited a range of responses. On Phase c, white pine rose from 6 to 10 m2/ha at year 110, and declined to 6 m2/ha by the end of the run. White pine persisted at low levels on Phases f and e. Red pine showed a general decline in basal area on all phases, as did paper birch. Balsam fir was present at low levels (1.5-2.5 m2/ha) in all phases. Trembling aspen showed a strong pulse on Phase f, reaching 10 m2/ha at year 30; this response did not occur on Phases c or e. The dominant response across all phases was the development of white spruce as a codominant, accounting for 36%, 48%, and 48% of basal area on Phases c, f, and e, respectively. Black ash, a dominant component of some of the plots in Phase e, slowly declined throughout the run. Also, in the more mesic conditions of phase e, sugar maple, not present in the initial sample, seeded in at year 10 and steadily increased to 4.2 m2/ha by the end of the run.
Soil nitrogen was consistently higher on the Guthrie Till Plain than on the Bena Dunes (Fig. 6). Available nitrogen exhibited a slow decrease during the course of the run in both Landtype Associations. In LTA E, nitrogen levels were generally higher in Phase i, although these values converged at year160. In LTA O, available nitrogen was lower in Phase c throughout most of the run.
Fig. 6. Simulated annual available nitrogen for six Ecological Landtype Phases, given current overstory starting conditions.
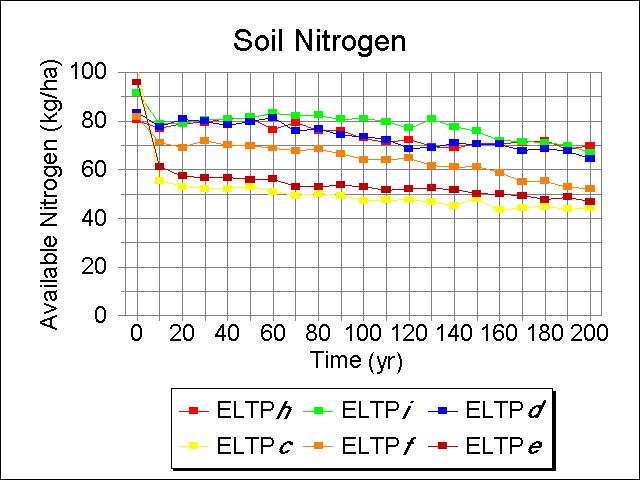
|
The temporal patterns of Foliage Height Diversity were fairly consistent across phases, increasing over the first 100 yr of the simulation, to a plateau that persists for the rest of the run (Fig. 7). Phase f was consistently half a unit lower than all other phases throughout the run. The lower structural diversity characteristic of this ecosystem implies that habitat is not as favorable for passerine songbirds (MacArthur 1965), particularly during the mid-successional phases of stand development.
Fig. 7. Temporal patterns of Foliage Height Diversity on six Ecological Landtype Phases, given current overstory starting conditions.
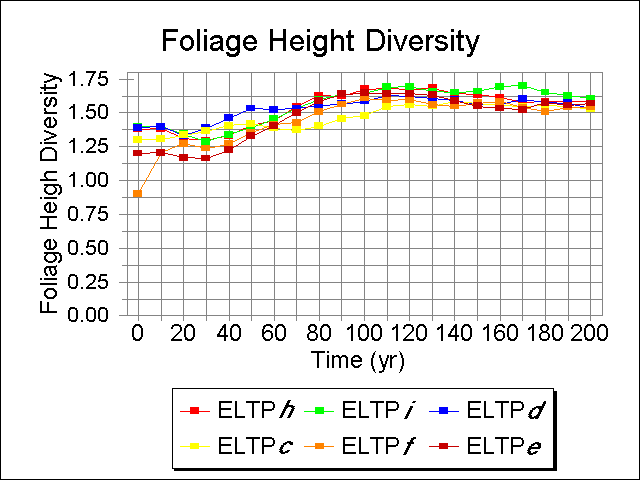
|
A fundamental tenet of ecological land classification is that the identified ecological units differ not only in state variables, but also in functional processes, such as nutrient cycling, forest succession, and response to management or disturbance. This ability to quantify spatial variation in ecosystem processes makes ecological classification an important tool for forest management, in that management strategies can be tailored to processes rather than states. To develop more sophisticated silvicultural treatments, however, requires an understanding of the dynamics of temporal processes. Simulation models provide a means of understanding these dynamics.
The LINKAGES model has had a number of applications, including assessing the response of northern forests to changing atmospheric CO2 levels (Pastor and Post 1988), predicting spruce decline (Pastor et al. 1987), and improving silvicultural practices (Pastor and Mladenoff 1992). In the present study, we parameterized the model for a set of ecological land units common in northern Minnesota forests. We then used the model to look at patterns of convergence and divergence in forest composition, given climatic, edaphic, and compositional characteristics of the map units. Units at the Phase level showed differences in future forest composition, although these differences were often expressed in terms of relative dominance rather than fundamentally different successional pathways. Yellow birch was found to be a major late-successional species in the northern hardwood-dominated morainal ecosystems, particularly Phases h and i, but was much less prevalent in the drier Phase d. Pine varied in its dominance on sandy landforms; this variation was due, in part, to limitations of soil moisture and to the long-term interactions of soil moisture status and nutrient quality. In the absence of disturbance, pine decreases in abundance as soil moisture increases.
Particularly important in this analysis is the fact that the variables mapped as part of the ecological classification are, in large part, the variables that drive ecosystem processes. In the hierarchical classification scheme developed by the Forest Service and implemented by numerous agencies (Cleland et al. 1997), a climatic classification provides overriding constraints on species range limits and potential productivity. Within climatic regions, glacial landform and local differences in soil, topography, and vegetation define fine-scale ecological units. Soil, in terms of moisture-holding capacity and nutrient capital, in conjunction with vegetation, which determines the potential rates of nutrient release, are the predominant driving factors. These local factors allow the simulation of phase-level nutrient dynamics, which, in turn, influences both potential composition and growth rates.
Although many critical factors in modeling forest growth are inherently part of the ecological classification system, others are not. Specifically, characterization of soil carbon and nitrogen levels is not part of the classification and mapping process, although numerous studies of the Lake States forests point out that these factors are fundamental drivers of ecosystem structure and function (Peet and Loucks 1977, Pregitzer et al. 1983, Pastor et al. 1984, Zak et al. 1986). The science of forest soils was historically derived from the perspective of the agronomic sciences, with an emphasis on pedogenesis and the description of texture, color, and horizonation of soils (Soil Survey Staff 1975). Soil taxonomy has also been limited to the upper soil horizons (the control section), and only weakly considers the effects of deep soil properties. Recent work has shown, however, that pedogenic processes in soils can extend far below the control section (Richter and Markewitz 1995), and that deep soil properties can fundamentally impact forest productivity (Host et al. 1988). In an agronomic setting, farmers routinely conduct soil tests to assess the productivity of their land and direct management. In forestry, soil testing is quite a foreign concept; forest managers are reluctant to add this to the suite of analysis currently used to characterize a forest plot. We maintain, given the importance of litter quality and quantity in forests, that a simple and relatively inexpensive program of soil testing would yield far more knowledge on ecosystem behavior than a detailed description of soil morphology. As the ecological management of forests becomes a matter of agency policy, the incorporation of these nontraditional ecological driving variables into site characterization might become more acceptable.
The combination of ecological land classification and modeling has a number of practical applications for developing forest management plans. First, the consequences of different silvicultural practices can be tested. For example, Pastor and Mladenoff (1992) compared the ecological consequences of clear-cutting vs. partial cutting. They found that the use of partial cuts, in which a harvest of aspen and birch was coupled with thinning from below, produced a greater product mix and a more diverse forest canopy. They also found that the silvicultural results varied depending on the soil type. Because soils are an integral part of ecological classification, it follows that the model can be used not only for assessing successional pathways among ecosystems, but also for predicting their response to silvicultural treatment. The advantage of the modeling approach is that various silvicultural prescriptions, both in terms of cutting regimes and timing of stand entry, can be tested with relative ease once a model is parameterized. Those that fare favorably in the model runs can then be carried on to test operations in the field. The use of this parallel modeling/experimentation approach has been shown to be valuable in numerous forestry operations (Host et al. 1996).
The analyses shown here can readily be incorporated into ecological field guides. They provide a more quantitative basis than those efforts in which successional pathways are hypothesized based on observational interpretations of life history and anecdotal evidence. Moreover, because these models are designed to run in the microcomputer environment, they can easily be used by forest managers directly. In conjunction with a reasonable training program, simulation models such as LINKAGES, parameterized with data collected in the ecological classification and inventory process, represent an important decision support tool with real-time applications to ecosystem management.
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a comment, follow this link. To read comments already accepted, follow this link.
This study was funded by the Minnesota Department of Natural Resources. Jack Benedix and Dan Buchman are acknowledged for their assistance in field work and laboratory analyses. This is contribution number 237 of the Center for Water and the Environment.
Cleland, D. T., P. E. Avers, H. McNab, M. E. Jensen, R. G. Bailey, T. King, and W. E. Russell. 1997. National Hierarchy of Ecological Units. Pages 181-200 in M. S. Boyce and A. Haney, editors. Ecosystem management: applications for sustainable forest and wildlife resources. Yale University Press, New Haven, Connecticut, USA.
Hill, M. O. 1979. DECORANA - A FORTRAN program for detrended correspondence analysis and reciprocal averaging. Section of Ecology and Systematics, Cornell University, Ithaca, New York, USA.
Host, G. E., J. G. Isebrands, G. W. Theseira, J. R. Kiniry, and R. L. Graham. 1996. Temporal and spatial scaling from individual trees to plantations: a modeling strategy. Biomass and Bioenergy 11:233-243.
Host, G. E., P. L. Polzer, and D. T. Cleland. 1995. Regional climatic analysis of the northern Lake States. Natural Resources Research Institute, Duluth, Minnesota, USA. NRRI/TR-95/32:1-37.
Host, G. E., and K. S. Pregitzer. 1991. Ecological species groups for upland forest ecosystems of northwestern Lower Michigan. Forest Ecology and Management 43:87-102.
Host, G. E., K. S. Pregitzer, C. W. Ramm, J. B. Hart, and D. T. Cleland. 1987. Landform-mediated differences in successional pathways among upland forest ecosystems in northwestern Lower Michigan. Forest Science 33:445-457.
Host, G. E., K. S. Pregitzer, C. W. Ramm, D. P. Lusch, and D. T. Cleland. 1988. Variation in overstory biomass among glacial landforms and ecological land units in northwestern Lower Michigan. Canadian Journal of Forest Research18:659-668.
Host, G. E., C. W. Ramm, E. A. Padley, K. S. Pregitzer, J. B. Hart, and D. T. Cleland. 1993. Field sampling and data analysis methods for development of ecological land classifications: an application on the Manistee National Forest. North Central Forest Experiment Station General Technical Report NC-162:47
Johnson, L. B., G. E. Host, J. K. Jordan, and L. L. Rogers. 1991. Use of GIS for landscape design in natural resources management: habitat assessment and management for female black bear. Proceedings of GIS/LIS '91 507-517.
MacArthur, R. H. 1965. Patterns of species diversity. Biological Review 40:510-532.
Pastor, J., J. D. Aber, C. A. McClaugherty, and J. M. Melillo. 1984. Aboveground production and N and P cycling along a nitrogen mineralization gradient on Blackhawk Island, Wisconsin. Ecology 65:256-268.
Pastor, J., R. H. Gardner, V. H. Dale, and W. M. Post. 1987. Successional changes in nitrogen availability as a potential factor contributing to spruce declines in boreal North America. Canadian Journal of Forest Research 17:1394-1400.
Pastor, J., and D. J. Mladenoff. 1992. Modeling the effects of timber management on population dynamics, diversity, and ecosystem processes. Pages 16-29 in D. C. LeMaster and R. A. Sedjo, editors. Modeling Sustainable Forest Ecosystems. Forest Policy Center, Washington, D.C., USA.
Pastor, J., and W. M. Post. 1986. Influence of climate, soil moisture and succession on forest carbon and nitrogen cycles. Biogeochemistry 2:3-27.
Pastor, J., and W. M. Post. 1988. Response of northern forests to CO2-induced climate change. Nature 334:55-58.
Peet, R. K., and O. L. Loucks. 1977. A gradient analysis of southern Wisconsin forests. Ecology 58:485-499.
Pregitzer, K. S., B. V. Barnes, and G. D. Lemme. 1983. Relationship of topography to soils and vegetation in an Upper Michigan ecosystem. Soil Science Society of America Journal 47:117-123.
Richter, D. D., and D. Markewitz. 1995. How deep is soil? BioScience 45:600-609.
Shadis, D., J. C. Almendinger, D. Hanson, B. Hargrave, J. Barott, and C. Adams. 1995. Chippewa National Forest: technical key to ecological landtype phases, Bena Dunes and peatlands. Chippewa National Forest, Cass Lake, Minnesota, USA.
Soil Survey Staff. 1975. Soil taxonomy. Soil Conservation Service Publication AH-436, Washington, D. C., USA.
Zak, D. R., K. S. Pregitzer, and G. E. Host. 1986. Landscape variation in nitrogen mineralization and nitrification. Canadian Journal of Forest Research 16:1258-1263.
ZedX. 1995. Database description: Minnesota, Michigan, and Wisconsin. Hi-Rez Data Climatological Series. ZedX, Boalsburg, Pennsylvania, USA.
Address of Correspondent:
George Host
Natural Resources Research Institute
University of Minnesota - Duluth
5013 Miller Trunk Highway
Duluth, Minnesota 55811 USA
Phone: (218) 720-4264
Fax: (218) 720-9412
ghost@sage.nrri.umn.edu
*The copyright to this article passed from the Ecological Society of America to the Resilience Alliance on 1 January 2000.
![]()