|
|
|
Copyright ©1998 by The Resilience Alliance*
Frelich, L.E. and P.B. Reich. 1998. Disturbance severity and threshold responses in the boreal forest. Conservation Ecology [online] 2(2): 7. Available from the Internet. URL: http://www.consecol.org/vol2/iss2/art7/
A version of this article in which text, figures, tables, and appendices are separate files may be found by following this link.
Insight, part of Special Feature on Sustainability and Resilience in Boreal Regions Disturbance Severity and Threshold Responses in the Boreal Forest Lee E. Frelich and Peter B. Reich
University of Minnesota
- Abstract
- Introduction
- Conceptual Models of Forest Response to Changes in Severity
- Which Model to Use: a Minnesota Case Study
- Discussion
- Responses to this Article
- Literature Cited
This essay discusses three potential models relating disturbance severity to post-disturbance stand composition in the boreal forest: (1) continuous, where changes in disturbance severity cause a proportional and continuous change in stand composition; (2) discontinuous, where a threshold disturbance severity exists beyond which major changes in composition occur; and (3) the cusp, where thresholds exist and coexistence of two alternative compositional states is possible at the same disturbance severity. Any of the three models may be appropriate in different stands or different parts of the boreal forest. If the actual model does not correspond to that assumed by forest managers, then forest harvesting practices may cause unexpected sudden changes in forest composition. Disturbance severity under the natural disturbance regime changes so dramatically, from one disturbance to the next, that oscillations in composition over time are likely to be individualistic and irregular, rather than stable. Harvesting operations are mainly done during winter, resulting in a very low disturbance severity in comparison to the natural regime, which included crown fires. Major changes in species compositon over large parts of the boreal forest are likely to result.
KEY WORDS: alternative stable states; disturbance in boreal forest; disturbance severity; forest response to changes in; Minnesota boreal forest; stability in boreal forest; forest response to changes in disturbance severity.
Many forest management models, which are used to project future forest composition and timber volume by species, assume that forest succession is a static process. That is, forests repeat the same regeneration and successional stages after each disturbance. This is based on the notion of resilience to disturbance (Holling 1973), and oscillation or stable cycles (Ludwig et al. 1997). However, in the case of the boreal forest, two factors that affect the successional trajectory of stands change constantly over time. These are the rotation period for natural disturbances (synonymous with fire cycle; Van Wagner 1978), and the severity of disturbances (degree to which trees and their propagules are killed, which we shall define in detail). The former factor is discussed by Bergeron (1998), whereas this paper covers the latter factor. We hypothesize that changes in severity of disturbances may lead to an individualistic successional pathway over time, rather than any sort of stable cycle.
There is budding consensus among forest ecologists that one of the best ways to preserve biodiversity in managed forests is to mimic the natural disturbance regime. Several authors have examined how the size of disturbances and rotation periods influence the successional trajectory. Bergeron (1998) questions the traditional use of the rotation period for severe disturbances, pointing out that it is always changing and, therefore, that the proportions of stands across the landscape that are early-, middle-, and late-successional are always changing. Severity also changes over time, and it is important to examine this aspect of the disturbance regime.
Along the southern edge of the boreal forest in eastern North America, pioneer species such as aspen (Populus tremuloides) and jack pine (Pinus banksiana) may dominate the landscape when frequent crown fires occur. However, during periods when crown fires have long rotation periods, many stands succeed to northern white cedar (Thuja occidentalis), often mixed with balsam fir (Abies balsamea), black spruce (Picea mariana), and paper birch (Betula papyrifera) (Buell and Niering 1957, Grigal and Ohman 1975, Bergeron and Dubuc 1989). The story here may not be as simple as long rotation periods; when crown fires are rare, other, ususally less severe disturbances, such as treefall and insect defoliation, become the most important feature of the disturbance regime. In fact, during those times when stand-killing fires are rare, disturbances of some sort still occur frequently.
Before examining the topic of disturbance severity in more detail, a definition is necessary. We propose a working definition similar to that of Oliver and Larson (1990), which equates severity with the proportion of individual trees and their propagules killed in a disturbance. Low-severity disturbance kills only a few scattered trees, either in the overstory or understory, leaving an esentially intact forest with a few gaps (e.g., treefall disturbance in old-growth forests). Moderate-severity disturbances kill most of the overstory or most of the understory, leaving either a canopy layer or seedling/seedbank layer intact (e.g., winter clear-cut logging, surface fire, or patchy crown fire). High-severity disturbances kill most of both the overstory and understory, leaving only a few scattered survivors and portions of the seedbank (e.g., intense crown fire). Very-high-severity disturbances remove virtually all individuals, including the seedbank. The very-high-severity disturbances, such as glaciation, landslide, and deposition of volcanic ash, are beyond the scope of this paper.
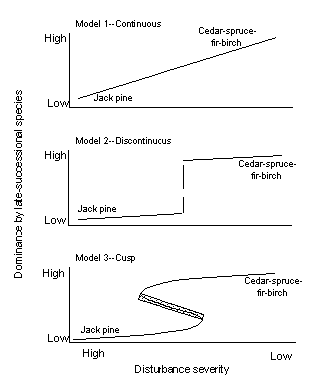
Changes in disturbance severity must necessarily cause changes in stand composition at some point. It is impossible for any one species in boreal forests of our region to be able to replace itself abundantly after disturbances spanning the range of severities discussed in this paper (intense crown fires to single treefall gaps). Instead, a given landform supports a successional complex of species wherein a shade-intolerant, early successional species group reproduces after severe disturbances, and a more shade-tolerant, late-successional species group reproduces after low-severity disturbance (Heinselman 1973, Grigal and Ohmann 1975, Frelich and Reich 1995). The main question in this paper is: what is the nature of the relationship between stand composition (dominance by early- vs. late-successional species) and disturbance severity? We propose three possible models for this relationship (Fig. 1). Keep in mind that the models show equilibrial attractors, like most regression-defined relationships in community or ecosystem ecology, and do not specify the route(s) or length of time by which a given stand will arrive at a given point. In other words, there may be lags after disturbance, or a change in disturbance regime, before a given stand approaches the line.
Fig. 1. Proposed models for the relationship between disturbance severity and forest tree species composition. Note that in the cusp model, sudden change would occur for any disturbance with severity beyond the edge of the cusp, in which case, composition would jump downward or upward. The shaded part of the curve is inaccesible.
|
Model 1. Continuous change.
Change in disturbance severity leads to a proportional change in species composition. Increasing or decreasing the severity of disturbance over time will lead to opposite pathways down or up the same slope.
Model 2. Discontinuous change without hysteresis.
Changing severity of disturbances will lead to very little change until a threshold is reached, causing a sudden jump in composition if severity is increased or decreased a tiny amount at the threshold. If disturbance severity is changed in the opposite direction, exactly the same pathway is used (i.e., no hysteresis).
Model 3. Cusp or discontinuous change with hysteresis.
Holling (1981) was one of the first to propose this model for use with forests and disturbance, and Ludwig et al. (1997) provide a recent synthesis. Coexistence of two forest types with different composition is possible with moderate severity disturbance, depending on the history of each stand. The changes that occur when moving from low to high severity disturbance are not the same as the opposite change in severity (i.e., hysteresis); the sudden jump in composition occurs at the edge of the cusp in each case, which is at a different point along the severity axis.The frequency of disturbance is also related to severity: if two disturbances come at time intervals so close that the system does not recover between the disturbances, then the severities of the two disturbances may be totally or partially additive. Heinselman (1973) gives a classic example. If two crown fires occur 50 years apart in a jack pine forest, there are plenty of seeds stored in serotinous cones to regenerate jack pine after the second fire. If the two fires are only 10 years apart, the second fire can leave the forest with few or no jack pine seeds, so that the forest will succeed to another species, usually aspen, in the case of northern Minnesota. Thus, two fires can literally have an additive effect on removal of jack pine individuals and propagules.
What type of evidence is necessary to judge which of these three models applies? Even more important, is there some type of evidence that forest planners and managers can aquire and use on a widespread basis? One way to tell which of the three models applies would be to have a series of experiments in which forests are subjected to disturbances of different severities, and then the response is observed. For example, an experiment could be done by dividing large stands of jack pine and of spruce - fir - birch into blocks, each subjected to a different disturbance severity, possibly including selection cuts, seed tree cuts, winter clearcuts, summer clearcuts, surface fire, passive crown fire, and intense crown fire. Immediately after the disturbances, the proportions of canopy trees, saplings, seedlings, and seeds in the seedbank could be counted and compared to the pre-disturbance counts, thus quantifying the disturbance severity. After observing the recovery of the forest for each treatment, an assessment could be made as to the successional trajectory, and the mature post-disturbance stand composition could be plotted along an axis from low to high dominance by late-successional species vs. observed disturbance severity. It would then be easy to see which of the three proposed models fits the data. Because trees are relatively long-lived, it would be necessary to follow post-disturbance composition for a few decades before we could be sure of the response. Unfortunately, data sets with long-term observation that also carefully recorded the disturbance effects on populations of trees and their propagules, and that also included a wide range of disturbance types and severities, are non-existent at the current time. However, some retrospective studies in which the disturbance regime has been well documented can partially illuminate the situation.
We can look at our study area in the southern boreal forest of northern Minnesota for a case study. The area is on the Canadian shield of granitic bedrock, with shallow soils and many lakes. It is well known that jack pine and aspen dominate the canopy of stands subjected to high-severity crown fires, whereas northern white cedar, black spruce, balsam fir (Abies balsamea), and paper birch (Betula papyrifera) dominate old-growth stands that have been under a low-severity disturbance regime of windfalls (single tree or small clumps of trees), and spruce budworm (Choristoneura fumiferana) infestation for at least two centuries (Heinselman 1973, Frelich and Reich 1995). This information merely tells us that there are stable forest types at each end of the disturbance-severity gradient, with no information on the shape of the curve. If jack pine - aspen forests and cedar - spruce - fir - birch forests can both perpetuate themselves after moderately severe disturbance, with little or no intrusion of species from the opposite group, then model 1 can be rejected. The coexistence of two forest types after each receives a disturbance of precisely the same severity (within the moderate-severity range) would be evidence for model 3. In practice, though, it may be impossible to measure the true severity of disturbance, especially taking into account the effects of disturbance on the seedbank, accurately enough to distinguish between models 2 and 3. If one were to observe a jack pine - aspen forest perpetuated after a surface fire, and a cedar - spruce - fir - birch forest perpetuated after clear-cut logging (both disturbances defined as moderate), it would be difficult to tell whether there was a small difference in the severity of the two disturbances, such that surface fire was just to the left of the discontinuity in model 2, and logging was just to the right of the discontinuity (Fig. 1). The field of disturbance ecology is not advanced enough to allow a timely measurement of true severity, other than general categories. Thus, relatively strong evidence of coexistence is necessary to prove the existence of the cusp in model 3 (Fig. 1). We suggest that perpetuation of two forest types by two different disturbance types that are within the moderate-severity range would be sufficiently strong evidence.
Returning to our case study, it is evident that cedar - spruce - fir - birch forests and jack pine - aspen forests in Minnesota can both be perpetuated by one type of moderate-severity disturbance: surface fire (Ahlgren 1970, Heinselman 1973). Rising heat from a surface fire can release seeds from serotinous cones of jack pine, and surface fires in the cedar - spruce - fir - birch forest often do not kill adult trees, assuring continued deposition of their seeds on the forest floor after surface fire. The situation for regeneration after other disturbances of moderate severity is more complex. The cedar - spruce - fir - birch species group generally can replace itself after both clear-cut logging and windstorms unless heavy deer browsing interferes. The jack pine - aspen forests are sometimes, but certainly not always, perpetuated by logging (although there may be a proportional shift toward aspen), and windthrow generally accelerates succession in these forests to the cedar - spruce - fir - birch species group (Grigal and Ohmann 1975, Kurmis et al. 1986, Frelich and Reich 1995). Thus, model 3 (Fig. 1) probably applies to some jack pine - cedar successional complexes in northern Minnesota, because coexistence of two forest types could occur over a range of moderate-severity disturbance. In other cases, however, we are not sure which of model 2 or 3 applies. This is in contrast with white pine (Pinus strobus) - birch forests in Minnesota, where there is a proportional relationship between disturbance severity and post-disturbance composition, with birch being progressively favored by disturbances of increasing severity (Heinselman 1973), so that model 1 is more appropriate.
Management implications of forest response to disturbance severity are important for several reasons. Landscape-level forest growth and change models, which are used to project future conditions of a forested landscape and to estimate future yield of fiber for each species, make a variety of assumptions regarding the effect of disturbance on composition. Some assume that clear-cutting will always lead to a pioneer species (e.g., aspen), or that all stands will regenerate to the average species composition of all young stands (e.g., 10 - 20 years old) across the landscape, or that pre-disturbance stand size and age are the most important influence, or that post-harvest stand regeneration will always be a function of the pre-harvest species composition, unless planting is undertaken (e.g., Jaakko Pöyry Consulting 1992, Vanclay 1992, Peterson and Carson 1996, Ek et al. 1997). These assumptions may lead to inaccurate modeling of future conditions. If managers are not aware of the probable correct model for response to disturbances of differing severities, and of how harvests vary in severity, surprises in stand response to manipulation may result. Sometimes they will expect a change in composition and get little or none, and at other times they will observe large, but unexpected, changes in stands. The following sections discuss the rationale and some applications for this.
Severity of logging vs. historical fire regimeMost stands on the boreal-forest landscape today originated after high-severity disturbance: a combination of logging followed by burning of slash in secondary forests, or crown fire in primary forests. Disturbance severity under the current harvesting regime is very different from the historical regime. Much logging in the central part of the North American boreal forest is done during the winter. This leaves the forest floor and seedling layer intact: a moderate-severity disturbance according to our definition. Therefore, the result could be a forest with intermediate composition between the two endpoints, a mixture of jack pine, fir, birch, spruce, and cedar, if model 1 applies (Fig. 1). If model 2 applies, then either maintenance of jack pine or a switch to spruce, fir, birch, and cedar could occur, depending upon exactly where the discontinuity and the severity of the new harvesting regime lie along the disturbance severity axis of Fig. 1. If model 3 applies, then the result could be similar to that of model 2, but maintenance of jack pine would be somewhat more likely, because the cusp provides that a wider range of disturbance severities is consistent with jack pine regeneration.
Changing the severity of the disturbance regime can change the composition, even if rotation periods and sizes of disturbances are purposely made the same as in the natural disturbance regime. Numerous recent studies have shown that logging does not always mimic natural disturbance, precisely because of differences in severity of the disturbance (Ahlgren 1970, Noble et al. 1977, Abrams and Dickman 1982, Carleton and MacLellan 1994).
Disturbance severity is hard to characterize over the landscapeInventory systems and data are available that can measure species composition very well, at least in terms of basal area of mature trees. However, forest inventory data are not sufficient to measure disturbance severity across the landscape, even though disturbance severity and impact can be measured accurately on a few intensively studied research plots. The legacy of seeds and seedlings left after a disturbance is especially important for future stand development, but is especially hard to inventory across the landscape, and is dependent on the stage of stand development within each stand at the time of disturbance (Whelan 1995, Peterson and Carson 1996).
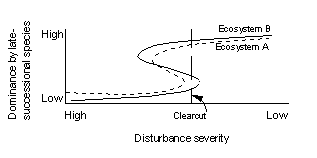
Any of the three models could be in effect for a given forest region. The appropriate model could differ over time with changes in stage of stand development, or could differ over space by ecosystem type. A hypothetical example (Fig. 2) shows how the location of the cusp in model 3 could vary, so that clear-cutting disturbance may not be severe enough to perpetuate a forest dominated by early-successional species in ecosystem type A, thus allowing, or even accelarating, succession to late-successional species (Abrams and Scott 1989). The same cutting, however, may be severe enough to perpetuate a forest dominated by early-sucessional species in ecosystem type B. Such differences in response to disturbances of roughly equivalent severity could explain why clear-cutting of black spruce forests commonly causes replacement by aspen on the Ontario Clay Belt (Carleton and MacLellan 1994), but the replacement by aspen does not necessarily occur in northern Minnesota, a different ecosystem type with poor soils (Grigal and Ohmann 1975).
Fig. 2. Hypothetical example showing how differences in the disturbance severity - composition relationship could influence forest response to a clearcut. If a stand currently dominated by early-successional species is clear-cut, two different responses could occur. For ecosystem A, the disturbance is lower than the threshold disturbance severity necessary to maintain dominance by early-successional species, and the system would shift to high dominance by late-successional species after the harvest. For ecosystem B, the same harvest would allow early-successional species to be maintained. For either ecosystem, clear-cutting in a stand currently dominated by late-successional species (on upper part of curve), would not cause any significant change in composition.
|
Stable oscillation vs. individualistic successional pathways
How likely is the repeated occurrence of disturbances similar in severity over time in one forest stand? The current literature suggests that, at least in boreal forests, such a sequence is unlikely. In boreal forests, forest fire intensity, or the rate at which heat energy is released per unit time, is very dependent on the degree of dryness of fuels and the weather at the time of the fire (Johnson 1992). Fire intensity has a large influence on whether a fire will stay on the ground or enter the crowns within a closed-canopy conifer forest, and this in turn determines whether the canopy layer is killed. Thus, fire severity is extremely variable from one event to the next: literally as variable as day-to-day weather. Even surface fires and prescribed fires vary greatly in severity (Whelan 1995). Windstorms have a similar variability in degree of blowdown caused, depending on windspeed and condition of the stands hit by the wind (Frelich and Lorimer 1991). Budworm damage in the boreal forest depends on stand composition at both the stand and landscape levels (Bergeron et al. 1995), thus leading to a wide range of tree mortality among stands. In addition, the timing of natural disturbances in the boreal forest is very erratic in a given stand. For example, high-severity crown fires in several study areas have been shown to burn stands of all ages with equal chance (VanWagner 1978, Johnson 1992). The chance of stand-killing fire does not increase with age, so that the cycle does not necessarily have to go back to the early-successional stage within a certain time frame.
Given this wide variability in the severity of sequential disturbance, forest types that follow model 1 will always be undergoing compositional change, whereas models 2 and 3 provide some buffering of forest response to changing severity, although the trade-off is that sudden, large changes can occur. Ludwig et al. (1997) describe parts of the boreal forest where preferential browsing of aspen and birch in young stands and poor reproduction of aspen under its own canopy lead to succession to fir, which in turn increases the density of fir foliage in the canopy, until budworm has an ideal breeding environment, and an outbreak occurs that kills much of the canopy. Then fire occurs and the system goes back to the aspen - birch stage. There are no such feedback factors, such as the response of budworm to foliage density in Ludwig et al. (1997), that would cause a regular oscillation with regard to disturbance severity in the boreal forest. The severity of each in a sequence of successive disturbances suffered by one stand is essentially independent of the last. Thus, in cases in which disturbance severity is the main driver of compositional change, stable oscillation is unlikely. In fact, the paleoecological analyses of Swain (1973) and Bergeron (1998) show that climate change, by regulating the disturbance regime, has governed the balance between the early-successional species group (jack pine - aspen) and the late-successional species group (white cedar - spruce - fir - birch) over the last several millenia in the southern part of the boreal forest.
Spatial structure of the forest at several scales also influences the chances of a regularly oscillating system. If the forest in northern Minnesota is to switch back and forth from jack pine to cedar as the disturbance regime changes, the seed source for species in each forest type must always be present. However, cedar and fir are often removed from stands by severe fires (Frelich and Reich 1995), and the placement of refuges from fire and the size of fire cause large variation in the amount of time it takes for cedar and fir reinvasion after fire. This could vary from a few years to centuries. Because of this, as well as climate changes, the forest is not likely to cycle through the jack pine and cedar phases on any regular basis. Instead, the successional sequence is probably individualistic over time, i.e., a given stand's trajectory may continuously vary, depending on the sequence of disturbances it receives.
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a comment, follow this link. To read comments already accepted, follow this link.
Abrams, M. D., and D. E. Dickman. 1982. Early revegetation of clearcut and burned jackpine sites in northern Lower Michigan. Canadian Journal of Botany 60: 946-954.
Abrams, M. D., and M. L. Scott. 1989. Disturbance-mediated accelerated succession in two Michigan forest types. Forest Science 35: 42-49.
Ahlgren, C. E. 1970. Some effects of prescribed burning on jack pine reproduction in northeastern Minnesota. University of Minnesota Agricultural Experiment Station. Miscellaneous Report 94, Forestry Series 5-1970.
Bergeron, Y., P.J.H. Richard, C. Carcaillet, S. Gauthier,
M. Flannigan, and Y.T. Prairie. 1998. Variability in fire
frequency and forest composition in Canada's southeastern boreal
forest: a challenge for sustainable forest management. Conservation
Ecology [on line] 2(2):6 . Available from the
Internet. URL: http://www.consecol.org/vol2/iss2/art6
Bergeron, Y., and M. Dubuc. 1989. Succession in the southern part of the Canadian boreal forest. Vegetatio 79: 51-63.
Bergeron Y., A. Leduc, H. Morin, and C. Joyal. 1995. Balsam fir mortality following the last spruce budworm outbreak in northwestern Quebec. Canadian Journal of Forest Research 25: 1375-1384.
Buell, M. F., and W. A. Niering. 1957. Fir - spruce - birch forest in northern Minnesota. Ecology 38: 602-610.
Carleton, T. J., and P. MacLellan. 1994. Woody responses to fire versus clear-cutting logging: a comparative survey in the central Canadian boreal forest. Ecoscience 1:141-152.
Ek, A. R., A. P. Robinson, P. R. Radtke, and D. K. Walters. 1997. Development and testing of regeneration imputation models for forests in Minnesota. Forest Ecology and Management 94:129-140.
Frelich, L. E., and C. G. Lorimer. 1991. Natural disturbance regimes in hemlock - hardwood forests of the Upper Great Lakes Region. Ecological Monographs 61:145-164.
Frelich, L. E., and P. B. Reich. 1995. Spatial patterns and succession in a Minnesota southern boreal forest. Ecological Monographs 65: 325-346.
Grigal, D. F., and L. F. Ohmann. 1975. Classification, description, and dynamics of upland plant communities within a Minnesota Wilderness Area. Ecological Monographs 45: 389-407.
Heinselman, M. L. 1973. Fire in the virgin forests of the Boundary Waters Canoe Area, Minnesota. Quaternary Research 3: 329-382.
Holling, C. S. 1973. Resilience and stability of ecological systems. Annual Review of Ecology and Systematics 4: 1-23.
_______ . 1981. Forest insects, forest fires, and resilience. Fire regimes and ecosystem properties. USDA Forest Service General Technical Report WO-26: 445-464.
Jaakko Pöyry Consulting. 1992. Maintaining productivity and the forest resource base. A technical paper for the Generic Environmental Impact Statement on Timber Harvesting and Forest Management in Minnesota. Jaakko Pöyry Consulting, Tarrytown, New York, USA.
Johnson, E. A. 1992. Fire and vegetation dynamics: studies from the North American boreal forest. Cambridge University Press, Cambridge, UK.
Kurmis, V., S. L. Webb, and L. C. Merriam. 1986. Plant communities of Voyageurs National Park, Minnesota, USA. Canadian Journal of Botany 64: 531-540.
Ludwig, D., B. Walker, and C. S. Holling. 1997. Sustainability,
stability, and resilience. Conservation Ecology [online]
1 (1): 7. Available from the Internet. URL: http://www.consecol.org/vol1/iss1/art7
Noble, M. G., L. K. DeBoer, K. L. Johnson, B. A. Coffin, L. G Fellows, and N. A. Cristensen. 1977. Quantitative relationships among some Pinus banksiana - Picea mariana forests subjected to wildfire and postlogging treatments. Canadian Journal of Forest Research 7: 368-377.
Oliver, C. D., and B. C. Larson. 1990. Forest stand dynamics. McGraw-Hill, New York, New York, USA.
Peterson, C. J., and W. P. Carson. 1996. Generalizing forest regeneration models: the dependence of propagule availability on disturbance history and stand size. Canadian Journal of Forest Research 26: 45-52.
Swain, A. M. 1973. A history of fire and vegetation in northeastern Minnesota as recorded in lake sediments. Quaternary Research 3: 383-396.
Vanclay, J. K. 1992. Modelling regeneration and recruitment in a tropical rain forest. Canadian Journal of Forest Research 22: 1235-1248.
Van Wagner, C. E. 1978. Age-class distribution and the forest fire cycle. Canadian Journal of Forest Research 8:220-227.
Whelan, R. J. 1995. The ecology of fire. Cambridge University Press, Cambridge, UK.
Address of Correspondent:
Lee E. Frelich
University of Minnesota, Department of Forest Resources
1530 N. Cleveland Ave.
St. Paul, Minnesota 55108 USA
Phone: 612-624-3671
Fax: 612-625-5212
freli001@maroon.tc.umn.edu
*The copyright to this article passed from the Ecological Society of America to the Resilience Alliance on 1 January 2000.
![]()