






The following is the established format for referencing this article:
Tito, R., N. Salinas, E. G. Cosio, T. E. Boza Espinoza, J. G. Muñiz, S. Aragón, A. Nina, and R. Roman-Cuesta. 2022. Secondary forests in Peru: differential provision of ecosystem services compared to other post-deforestation forest transitions. Ecology and Society 27(3):12.ABSTRACT
While tropical forests are undergoing rapid transformation as a result of direct human impacts, many deforested areas are reverting to forest through natural or human-assisted regeneration. This situation provides a window of opportunity to implement forest management strategies to achieve environmental objectives while promoting social development and contributing to local livelihoods. Successful forest management policy, however, depends on how well we can appraise environmental consequences as well as on the value of ecosystem services that these regrowing forests provide. Here, we review the published literature to synthesize the ecosystem services provided by three types of forest transitions: naturally-regenerated secondary forests, agroforestry systems, and tree plantations, in the coastal, Andean, and Amazonian regions of Peru. We then discuss the potential of these regrowing forests as nature-based solutions that can help in the adoption of policies that promote their sustainable use and conservation. Our literature analysis reveals that forest transitions provide significant services in offsetting carbon emissions, providing habitats for biodiversity, and regulating hydrological services. However, the amount and importance of ecosystem services vary depending on the forest transition type. Secondary forests offer multiple services, representing a low-cost, immediate, and highly effective strategy in mitigating the climate and biodiversity crises and ultimately providing vital ecosystem services to society, such as water provision. In contrast, exotic tree plantations have negative effects on water regulation services. We highlight the potential of secondary forests for land management that supports multiple and integrated environmental initiatives. This framework can guide policy decisions to choose appropriate options on forest transition types most suitable to achieve specific end goals at local and regional scales, considering both ecosystem services and disservices to avoid trade-offs in which the achievement of one goal is detrimental to another.INTRODUCTION
Tropical mature forests store large amounts of carbon and host high biodiversity, but they are undergoing rapid transformation as a result of direct human impacts and changing environmental conditions (Hansen et al. 2013, Colorado Zuluaga and Rodewald 2015, FAO 2020). This context of forest degradation in Peru is not much different from that in other countries. In its recent report on deforestation, the Peruvian National Forest and Wildlife Service (SERFOR) revealed that between 2017 and 2019, the rate of forest loss in the Peruvian Amazon averaged 128,069 ha/yr and had increased compared to the previous 10 years (https://geo.serfor.gob.pe/visor/). Similarly, other studies have shown increasing forest disturbance rates in many areas of the Andes and coastal regions (Aide et al. 2019, FAO 2020). The main causes of forest loss are related to the expansion of agriculture and pasture in response to growing demand for food and other basic products. In addition, deforestation by gold mining is also responsible for a significant extent of forest loss (Tovar et al. 2013, Caballero Espejo et al. 2018, Csillik and Asner 2020). Immediate and observable economic benefit provided by this change in land use outweighs, in the near term, the loss of ecological benefits from forests that are often intangible to human perception (D’Almeida et al. 2007, Giam 2017, Moomaw et al. 2019).
Many areas deforested for timber extraction, agriculture, or pasture are reverting to forest through natural or human-assisted regeneration (i.e., by planting trees; Chazdon et al. 2020). These “new forests” or “reforests” that recover their forest cover in an increasing trend, commonly following a period of deforestation in line with socioeconomic changes, are known as “forest transitions” (Wilson et al. 2017, MacDonald and McKenney 2020). Naturally regenerating forests on abandoned deforested lands (i.e., secondary forests) are increasingly expanding in many regions and are likely to be a dominant feature of tropical forests in the near future (Poorter et al. 2016, Wilson et al. 2017). Recently, it has been estimated that secondary forests represent half of the remaining forest in tropical regions (McGee et al. 2020). Approximately 2.6–8 million ha have been reported as secondary vegetation in the Peruvian Amazon (MINAM 2015, Smith et al. 2021). Published reports have shown that secondary forests often rapidly accumulate aboveground biomass and thus sequester carbon dioxide at even higher rates than mature forests (Asner et al. 2010, Poorter et al. 2016, Chazdon et al. 2016). In addition to carbon sequestration, secondary forests also host high biological diversity and generate critical provisioning ecosystem services such as water, timber, or food (Poorter et al. 2016, Jones et al. 2019). Despite these and other important benefits, secondary forests are still undervalued, both ecologically and economically, and are therefore largely neglected in forest management policies.
In addition to secondary forests, tropical landscapes are also increasingly being occupied by agroforestry systems and tree plantations (Horgan 2009, Socolar et al. 2019). Although these sytems are not naturally regenerated forests, many studies advocate the potential for these human-managed tree-covered areas to deliver important services and benefits (Ehrenbergerová et al. 2016, Zavala et al. 2018, Jezeer et al. 2019). However, they are ecologically distinct from each other and from the original land cover (i.e., mature forest; Wilson et al. 2017), and the environmental consequences, as well as the value of the ecosystem services that they ultimately provide, are still poorly understood (Barbier et al. 2010, Wilson et al. 2017, MacDonald and McKenney 2020). For example, the expansion of certain types of forest plantations (e.g., exotic tree plantations) provides important provisioning and supporting services, but can also degrade other services such as water- and soil-related services (Raboin and Posner 2012, Vallet et al. 2016, Bonnesoeur et al. 2019). When the real benefits of forests are unknown, it also leads to unclear forest policy. In this sense, a simultaneous analysis of changes in forest cover type, structure, function, and services can provide valuable information for the design of appropriate land management strategies (MacDonald and McKenney 2020). Therefore, estimating the ecological and social benefits provided by distinct forest types is a necessary first step.
Given the growing awareness about the impacts of climate and land-cover change, a number of international and national initiatives have been proposed to protect intact forests, reduce deforestation, and restore degraded lands through natural or assisted forest regeneration (Shukla et al. 2019). The forest and landscape restoration agenda that seeks to limit global climate change by removing carbon dioxide from the atmosphere through the growth of trees is an example of such initiatives that are currently being adopted (Shukla et al. 2019, Chausson et al. 2020, Soto-Navarro et al. 2020). One such activity is the Bonn Challenge, an international commitment to restore 350 million ha by 2030 (Holl and Brancalion 2020; https://www.bonnchallenge.org/). Under Initiative 20x20 (Buenos Aires Declaration 2019: https://initiative20x20.org/news/buenos-aires-declaration-restoration), Peru has pledged to restore 3.2 million ha, targeting 2 million ha for commercial tree plantations and the remaining 1.2 million ha aimed at land under different degradation modalities (overgrazing, salinity, water erosion, soil pollution, and soil compaction; Román et al. 2018, Cerrón et al. 2019). However, these strategies face significant challenges. Afforestation and reforestation carry significant costs in time and money for both implementation and long-term monitoring. Indeed, long-term monitoring is often not in place, and short-term successes of afforestation and reforestation do not translate into durable and effective forest recovery (Cerrón et al. 2017, 2019, SERFOR 2018). In contrast, naturally regenerating secondary forests could be a more suitable approach to optimize the provision of multiple ecosystem services because these forests are recovering in areas where environmental conditions allow it, bypassing problems of water supply and changing climate that can affect the establishment of human-led afforestation and reforestation efforts on degraded land (Chazdon and Guariguata 2016). Deciding which forest transition types can provide multiple and effective services in the short term is of extreme importance in the context of finding effective tools for mitigation and adaptation to climate change and in supporting integrated, sustainable, land-use management (Chazdon and Guariguata 2018).
To help improve decision-making on naturally regrowing forests and their management in Peruvian landscapes, we examined the available literature concerning different tree-based approaches for restoring degraded land. We focused only on forested transitions because the multiple services they provide are considered a win-win situation compared to other agricultural options under the current challenges of climate change, biodiversity loss, and desertification. Here, we analyze three different forest transitions (secondary forests, agroforestry systems, and tree plantations) and their provision of three key ecosystem services (carbon sequestration and storage, habitat for biodiversity, and water regulation) in Peru’s three main regions (coastal, Andean, and Amazonian). We also provide information on the ecosystem services offered by mature native forests to provide a reference state for comparison with those provided by these forest transitions. More specifically, we address the following key questions: What ecosystem services might we expect from secondary forests and other post-deforestation forest transitions? What is the relative importance of these systems in each region? And, how can this information help in designing effective social and environmental policies?
STUDY CONTEXT AND DATA SOURCES
Study context
Peru hosts an ample variety of ecological zones, with a total of 84 of the 104 ecological regions in the world, and 28 different climates (Escobal and Torero 2003, MINAM 2014). The heterogeneous habitats of Peruvian landscapes sustain high biodiversity and provide critical services to society. Approximately 22% of the Peruvian national economy is linked to biodiversity and the trade of biodiversity products, which represents > $200 million USD (MINAM 2014). At the same time, Peru is identified as the world’s most vulnerable country to climate change and is subject to high pressure of land-use change (MINAM 2015, 2016, Gobierno del Perú 2020). Thus, because of its varied geographical characteristics and nature-dependent economic activities, Peru faces serious challenges from the effects of global change. In this context, forests will play crucial roles in the efforts to preserve biodiversity and mitigate climate change and its associated effects.
Amazonian forests: Peru’s vegetation covers 103 million ha, with 72 million ha corresponding to forests, mostly in lowland Amazon (MINAM 2015) below 500 m elevation (Fig. 1A). This region is the least populated area of Peru but is responsible for the greatest amount of forest extraction. As of 2017, 17% of the original forest area in the total Amazon basin had been cleared (Bullock et al. 2020), and 6.9% in the Peruvian amazon (Smith et al. 2021). The average annual forest loss in the Peruvian lowland Amazon forest was 128,764 ha/yr for the period 2000–2020 (FAO 2020). Agriculture and gold mining are expanding at an unprecedented rate in the Amazon, and these activities are the main trigger of deforestation in Peru (Caballero Espejo et al. 2018, Manoli et al. 2018, Csillik and Asner 2020). For example, the expansion of oil palm plantations in the last two decades in the northeastern Peruvian Amazon increased dramatically, from 15,000 ha in 2000 to > 108,000 ha in 2019, resulting in 2.8 Pg of carbon emissions (Escobedo Grandez 2021). Oil palm plantations account for approximately 11% of the deforestation from agricultural expansion from 2007–2013 (Vijay et al. 2018). In contrast, gold mining was responsible for 1.12 Tg/yr of carbon emissions (Csillik and Asner 2020), and its influence is largely growing in Peru, particularly in Madre de Dios (Caballero Espejo et al. 2018, Álvarez-Berríos et al. 2021).
Andean forests: The Andean region (27% of the country) comprises a longitudinal mountain range that crosses the region from north to south (Fig. 1A). It ranges from 2000 m above sea level in the dry western slopes and 500 m above sea level in the humid eastern slopes to alpine regions with glaciers > 6000 m above sea level. With 12.2 million ha in 2011, forests in the Andes represent ~21% of the total forested land in the country (MINAM 2015, 2016). These forests hold a disproportionate importance in providing hydrological services and supporting biodiversity, but rapid land-use change poses an increased threat to both biodiversity and ecosystem services availability. Thus, coffee, cocoa, and tea plantations (Fig. 1B–D), as well as fire and afforestation and reforestation with exotic tree species, are increasing the pressure on Andean natural land cover (Tovar et al. 2013, FAO 2016, Oliveras et al. 2018). In 2018, shade coffee agroforestry occupied approximately 5% (608,332 ha) of the total extent of the Andes (FAO 2020). By 2012, > 1 million ha of Andean nonforested areas (natural grasslands and degraded lands) were reforested with plantations (FAO 2016). Most of these plantations have tended to focus on exotic Eucalyptus and Pinus species because of their fast growth and economic profitability in short periods (FAO 2016, Cerrón et al. 2017, SERFOR 2018). In the Cajamarca region, for example, Eucalyptus and Pinus plantations replacing natural grasslands increased by 12.3%/yr during the period 1987 to 2007 (Tovar et al. 2013). In addition to human impacts, climate is rapidly changing in the Andes, with rising temperatures that lead to glacier melt, altered weather patterns, and associated long-term droughts (Vuille et al. 2008, 2018).
Dry coastal region and seasonally deciduous forests: Covering 11.7% of Peru’s total area, this region is a narrow longitudinal strip that extends from the Pacific Ocean to 2000 m above sea level in the dry western Andean slopes (SERFOR 2019; Fig. 1A). Despite its water constraints, this region is home to ~55% of the human population (MINAM 2016), with the Peruvian capital, Lima, settled in a western coastal desert. Although few studies have estimated the rate of dry forest deforestation in Latin America and Peru, deforestation is indeed a serious problem in Peruvian coastal forests (Whaley et al. 2010, Pécastaing and Chávez 2020). In the Piura region, for instance, it was estimated that between 15,000 and 40,000 ha of dry forest are lost annually (Pécastaing and Chávez 2020). Urban demand for firewood and charcoal, combined with agriculture and livestock expansion, are the main threats to the conservation of dry forests (Whaley et al. 2010, Bennett-Curry et al. 2013, SERFOR 2019). These human activities potentiate impacts from enhanced climate variability, such as El Niño–Southern Oscillation-related events (Pécastaing and Chávez 2020), which, in northern Peru, manifest in the form of increased coastal precipitation and flooding (El Niño-Costero), and in the south, as severe drought (Bourrel et al. 2015).
Data sources
We ran the search string for both English and Spanish publications in the Google Scholar database up to September 2020 and conducted a review and analysis of published peer-reviewed articles, master’s dissertations, doctoral theses, and research reports on the topic of ecosystem services for our three selected forest transitions: secondary forest, tree plantations, and agroforestry systems. In the tree plantation category, we included commercial, monoculture, and exotic tree plantations. In the agroforestry systems category, we included tree intercropping systems, tree-shaded perennial systems, windbreaks, and live fence systems. We focused on ecosystem services that are essential to nature and human well-being, considering current and future environmental change conditions. Thus, we reviewed publications that assessed at least one of the following three ecosystem services: carbon stocks and sequestration, habitat for biodiversity, and water regulation. We used a combination of different key words (“secondary forests”, “second ground forests”, “agroforestry”, “tree plantation”, “natural regeneration”, etc.) to identify literature related to our research interests. Despite initially focusing on peer-reviewed journal articles, we extended our literature search to “gray literature” (i.e., master’s or doctoral theses, research reports) to identify all available data related to the review topic. Because of the large number of papers retrieved in the search, we performed a first round of screening based on a review of titles and abstracts to select the articles most related to our goals. A total of 163 abstracts were initially identified as meeting the inclusion criteria, but further screening resulted in a total of 76 papers that were included in this review. Only studies conducted in Peru were considered.
RESULTS AND DISCUSSION
Forest ecosystem services
In forest transition theory (i.e., switch from deforestation to increasing forest cover), forests are often simply defined as forest cover and, an increase in forest cover is assumed to improve ecosystem services (Barbier et al. 2010, Garcia et al. 2020). However, forests deliver unique sets of services with varying quality and quantity depending upon their type, characteristics, and environmental conditions. This situation occurs because different pathways and drivers of forest recovery combined with different ecological characteristics, distributions, and trajectories lead to different suites of ecosystem functions and forest services (Barbier et al. 2010, Vallet et al. 2017, Wilson et al. 2017, Garcia et al. 2020, MacDonald and McKenney 2020).
Through the literature analyses, we found empirical evidence showing that regenerating forests provide important carbon sequestration sinks and habitats for a wide taxonomic range of wildlife (Box 1–3, Figs. 2 and 3). Forest cover recovery also has positive or negative effects on hydrology, depending on specific characteristics of the forest type and region (Box 1–3). As expected, the amounts and values of ecosystem services provided by secondary forests, agroforestry systems, and tree plantations vary among the forest types and regions.
Amazonian forest ecosystem services
Amazonian forests are the largest remaining terrestrial carbon stock (e.g., Saatchi et al. 2007, 2011, Asner et al. 2010). Peruvian lowland Amazon forests, in particular, have the potential to store > 150 Mg/ha of carbon in their aboveground biomass (Asner et al. 2014). Across the Peruvian Amazon, a recent study reported 2.6 million ha of secondary forest, and this forest recovery resulted in an accumulation of 73.8 Tg of carbon (Smith et al. 2021).
Our literature review reveals that Amazonian secondary forests are promising pathways to mitigating climate warming while protecting biodiversity (Box 1, Fig. 2A). Higher productivity in secondary forests suggests up to 11 times faster growth and carbon uptake in secondary forests than in mature forests (Poorter et al. 2016). Recent research, however, suggests that this enhanced growth is severely affected by repeated droughts, and secondary forests’ uptake can be overestimated (Smith et al. 2020). Compared to other forest pathways, studies suggest that carbon stocks in secondary forests recover at a faster rate than in tree plantations and agroforestry systems (Box 1, Fig. 2A). Thirty-year-old secondary forests in the Peruvian Amazon stored 50–60% of mature forest values (Málaga et al. 2021), and 50-year-old secondary forests store comparable amounts of carbon to mature forest (Chuquizuta et al. 2016; Box 1, Fig. 2A). Although agroforestry systems and tree plantations store important quantities of carbon, they still result in high proportions of carbon losses compared to mature forest conversion, particularly when plantations are fast-growth, low wood-density exotic species (Box 1, Fig. 2A). These findings support the claims made by previous studies (Wilson et al. 2017, Lewis et al. 2019).
In addition to the carbon sequestration potential, secondary forests harbor a substantial amount of biodiversity (Box 1, Fig. 2B). For example, 30- to 50-year-old secondary forests host 80–100% of the bird, mammal, and reptile species occurring in nearby mature forests (Whitworth et al. 2016) and are especially important habitats for large-bodied animals (Tapirus terrestris, large primates, Priodontes maximus) and for several threatened species (e.g., Puma concolor; Gavin 2004). In contrast, agroforestry systems and tree plantations have much simpler structure than in secondary forests and provide habitats for smaller species (Box 1, Fig. 2B). The recovery of dung beetle and termite species was higher in agroforestry systems than in secondary forests, and a high proportion of insect species recovery was also reported in agroforestry systems and tree plantations (Box 1, Fig. 2B). Some compositional and structural characteristics of human-managed forests can improve the occurrence of some species. In the Brazilian Amazon, for example, high numbers of mature forest bat, epigeic arachnid, lizard, and dung beetle species were found in areas of exotic tree plantations with an understory of native shrubs, similar to those occurring in secondary forests (Barlow et al. 2007). The value of conserving the biodiversity is not only to maintain species and genetic diversity, but also for the benefits and services they bring to humans. Accordingly, many of these wild trees and animals occurring in naturally regenerating areas also provide important food sources and additional income for local inhabitants (Gavin 2004, 2007, Fitts et al. 2020).
Here, we summarize the main findings for three key ecosystem services provided by secondary forests and post-deforestation forest transitions (agroforestry systems and tree plantations) in the Amazonian region of Peru. For a complete list of studies reviewed and their carbon stock values and species occurrences see Appendix 1 and 2.
Carbon stocks
Mature forest
The reported aboveground carbon stock in the lowland Amazon ranges between 107.6 and 335.1 Mg/ha (Martel and Cairampoma 2012, Sosa Castillo 2016). Total carbon stock ranges from 155.6–632.4 Mg/ha (Barbarán 1998, Martel and Cairampoma 2012).
Secondary forest
Studies report that both aboveground and total carbon stocks increase with forest age (Fig. 2). For instance, aboveground carbon stocks in early (1–5 years old), young (15 years old), and old (50 years old) secondary forests are 2.4–42.1, 121–184.4, and ~282.8 Mg/ha, respectively (Alegre et al. 2003, Chuquizuta et al. 2016). Similarly, total carbon stock is 40.5–95.8, 153.3–239.1, and 396.8 Mg/ha in early, young, and old secondary forests, respectively (ICRAF 1998, Alegre et al. 2003, Chuquizuta et al. 2016). On average, 30-year-old secondary forests store 50–60% of the value of mature forests (Málaga et al. 2021), whereas 50-year-old secondary forests store similar amounts of carbon as mature forests (Chuquizuta et al. 2016).
Agroforestry systems
Many agroforestry systems in the lowland Amazon are Theobroma cacao or Coffea arabica shaded by two to six tree species. The most common species used in agroforestry are Inga edulis, Guazuna crinite, Calycophyllum spruceanum, C. spruceanum, Cedrela odorata, Mariosousa willardiana, Cajanus cajan, Schizolobium amazonicum, and Leucaena leucocephala. Aboveground carbon stock ranges from 2.9–4.4 Mg/ha in 1-year-old agroforestry systems to 27–32.4 Mg/ha in 20-year-old agroforestry systems, whereas total carbon content ranges from 33.4–37.2 to 168.9 Mg/ha in 1-year-old and 10-year-old agroforestry systems, respectively (Concha et al. 2007, Villogas Ventura 2013, Timoteo et al. 2016). However, the amount of carbon stock in agroforestry systems depends on the tree species, so these values may vary (Appendix 1).
Tree plantations
Aboveground carbon stocks in tree plantations vary from 9.5 Mg/ha in 1-year-old Guazuma crinita plantations (Baltazar Peña 2011) to 66.6 Mg/ha in 30-year-old Hevea brasiliensis plantations (Alegre et al. 2003). Total carbon stock is 152.3–152.6 Mg/ha in 30-year-old Hevea brasiliensis plantations (ICRAF 1998, Alegre et al. 2003).
While not forest, 6- and 30-year-old Bactris gasipaes plantations were reported to contain 72.2 and 90.9 Mg/ha of carbon in their total biomass, respectively (Chuquizuta et al. 2016, Cuellar Bautista and Salazar Hinostroza 2016).
Habitats for biodiversity maintenance
Secondary forest
Several studies showed high biodiversity in naturally regenerating forest, especially in old secondary forests; 30–50-year-old secondary forests host 73–95% of amphibian, 81–98% of bird, 79–81% of mammal, and 88–110% of reptile species found in nearby mature forest (Whitworth et al. 2016). In contrast, young secondary forests (9–15 years old) contain 74–83% of dung beetle, 36.8% of termite, and 47% of tree species found in surrounding mature forest (Horgan 2009, Culot et al. 2011, Gonzalez et al. 2014, Vebrova et al. 2014, Duran-Bautista et al. 2020; Fig. 2B, Appendix 2).
Agroforestry systems
In lowland Amazon, agroforestry systems contain a high diversity of dung beetles (87%) and insects (71–79%), but few tree (23–24%), termite (47%), and bird (14–16%) species (Horgan 2009, Lojka et al. 2011, Vebrova et al. 2014, Perry et al. 2016, Aerts et al. 2017, Duran-Bautista et al. 2020; Fig. 2B, Appendix 2).
Tree plantations
Tree plantations host less biodiversity than mature forests, secondary forests, and agroforestry systems. Forest tree plantations host 66.7% of insects and 30–39% of dung beetle species compared to those occurring in mature forest (Horgan 2009, Lojka et al. 2011, Aerts et al. 2017; Fig. 2B, Appendix 2).
While not true forest plantations, oil palm (Elaeis guineensis) plantations contain 60.5% of termite and 42.6% of bird species found in nearby mature forest (Srinivas and Koh 2016, Duran-Bautista et al. 2020).
Hydrological services
Water infiltration and soil moisture: While deforestation may decrease local precipitation, inducing drier soils and decreases in evapotranspiration (Marengo 2006, D’Almeida et al. 2007), natural regeneration by native tree species improves rainfall and infiltration and increases available moisture (Brandon 2014). Natural regeneration can rapidly (in 5–10 years) reverse the effects on water regulation (Hölscher et al. 2005). In cacao agroforestry systems, accumulation of plant litter produced by shade trees (Inga sp., Macrolobium acaciafolium, Calycophyllum spruceanum, Cedrelinga cateaniformes, and Vitex pseudolia) allows greater rainwater infiltration, preventing soil moisture loss and increasing soil water-holding capacity (Arévalo-Gardini et al. 2015). However, plant-available water is significantly lower in agroforestry soil (10.6–11.7%) than in 30-year-old secondary forest soil (12.3–13.4%; Arévalo-Gardini et al. 2015).
Water flow regulation and quality: Tropical mature forests provide a high degree of water infiltration with little erosion or surface runoff and improve water quality by preventing some sedimentation and erosion (Marengo 2006, D’Almeida et al. 2007, Brandon 2014). Amazon mature forests use more water and have higher evapotranspiration and infiltration than human-modified vegetation types (tree plantations or agriculture; Bruijnzeel 2004). Evapotranspiration is a key process in this humid Amazon region, representing 28% of total local inputs to precipitation (Ellison et al. 2012). Contrastingly, young tree plantations have high evapotranspiration (similar to mature forest) but lower infiltration, reducing groundwater recharge (Brandon 2014). Similarly, timber plantations replacing mature forests lead to increases in evaporative losses, resulting in decreases in annual stream flow (Scott et al. 2005).
Oil palm plantation expansion has boomed over the last decades, generating ecosystem degradation (Butler and Laurance 2009, Srinivas and Koh 2016). Young oil palm plantations significantly decrease ecosystem evapotranspiration (−40% without understory, −20% with ground cover) and infiltration rates, and strongly increase runoff (up to 40% without understory; Manoli et al. 2018). These changes in water flow cause hotter and drier local climate (i.e., changes in water yield). Nevertheless, mature tree plantations (> 8 years old) have higher gross primary productivity and transpire more water (up to 7.7% more water than the forests they replaced), and thus reduce water runoff (Manoli et al. 2018).
Deforestation in the lowland Amazonian region causes severe impacts on the local water cycle by decreasing local precipitation and increasing drought intensity (Marengo 2006, D’Almeida et al. 2007). Forest regrowth can reverse these effects by providing important services in local and regional hydrological processes, improving infiltration, rainfall, and moisture (Box 1). Studies have shown that Amazonian secondary forests improve the quality, yield, and delivery of fresh water at multiple scales, from groundwater to rivers and rainfall, and also sink atmospheric moisture and prevent floods (Box 1). Agroforestry also improves rainwater infiltration and thus increases water-holding capacity (Box 1). Tree plantations, in contrast, can have negative effects by increasing water costs and reducing infiltration, lowering groundwater recharge and annual stream flow (Box 1). For example, timber plantations (e.g., Eucalyptus, Pinus, Hevea) replacing old-growth forests lead to an increase in evaporative demand, resulting in a decrease in annual stream flow (Scott et al. 2005). In general, we highlight the potentially deleterious consequences on hydrology of the large-scale monospecific commercial plantations that are expanding in this high-biodiversity region with vital ecosystem services for society and the environment (Box 1).
Andean forest ecosystem services
The Andean region is exceptionally rich in biodiversity and has been highlighted as a hotspot of global biodiversity, with high endemism (Myers et al. 2000). It offers significant carbon storage in its soils, particularly water-logged ones (Román-Cuesta et al. 2011), and it is a fundamental regional water provider for both Andean slopes: the dry western Pacific coast (Beresford-Jones 2004, Whaley et al. 2010) and the eastern coast, whose Amazon river is sourced by Andean streams (Clark et al. 2014, Doornbos 2015).
Biomass in tropical Andean forests in Peru shows high variability related to variable environmental and topographic conditions. However, studies show that the aboveground biomass of the three transitions displays relatively similar values during the first decades of forest cover recovery, with tree plantations having the highest stocks (Box 2, Fig. 3). For example, ~10-year-old secondary forests, agroforestry systems, and tree plantations, respectively, contain on average 43, 54, and 72 Mg/ha of carbon in their aboveground biomass, representing approximately 20, 27, and 33% of the stock in mature forest (Box 2, Fig. 3). In the high Andes, cloud montane forests have low growth rates, and ~30-year-old secondary forests store only 32% of the mature forest carbon in their aboveground biomass (Oliveras et al. 2018). Contrastingly, these montane cloud forests can have relatively high rates of soil carbon accumulation due to the abundance of mosses (particularly Sphagnum) that store water and prevent litter decomposition (Oliveras et al. 2018). Soil restoration in secondary forests in the Andes can occur more rapidly than in other human-managed forest systems (Oliveras et al. 2018, Walentowski et al. 2018).
Here, we summarize the main findings for three key ecosystem services provided by secondary forests and post-deforestation forest transitions (agroforestry systems and tree plantations) in the Andean region of Peru. For a complete list of studies reviewed and their carbon stock values and species occurrences see Appendix 1 and 2.
Carbon stocks
Mature forest
The aboveground carbon stock in Peruvian montane forests is 52.4–485.3 Mg/ha (Lapeyre et al. 2004, Oliveras et al. 2018), and the total carbon stock is 154.3–398.8 Mg/ha (Chuquizuta te al. 2016, Oliveras et al. 2018).
In the high-elevation Andes, forest patches dominated by Polylepis and Escallonia species store lower amounts of carbon than other montane forests, ranging from 4.8–40.1 Mg/ha in aboveground biomass and from 23.8–148.7 Mg/ha of total carbon stock (Cuellar Bautista and Salazar Hinostroza 2016, Gurmendi Porras and Orihuela Izaguirre 2019).
Secondary forests
Aboveground carbon stock in early (1–5 years old) secondary forests is 26.1 Mg/ha, while young (~10 years old) and old (~30 years old) secondary forests store roughly equivalent amounts of carbon, at 43.2 and 36.6–42.3 Mg/ha, respectively. On average, ~30-year-old secondary forests store only 32% of carbon in aboveground biomass compared to mature forest (Oliveras et al. 2018, Aragón et al. 2021). Total carbon stock in these early, young, and old secondary forests increases with forest age (116.6, 161.1, and 295.3 Mg/ha carbon, respectively).
Agroforestry systems
Aboveground carbon stock in coffee agroforestry shaded by Inga, Pinus, or Eucalyptus ranges from 19.3–62 Mg/ha, while total carbon stock varies from 119.9–177.5 Mg/ha. Variation in carbon stocks are due to age and species used as shade (Lapeyre et al. 2004, Ehrenbergerová et al. 2016).
Tree plantations
Aboveground carbon stock increases with age in tree plantations, from 30.1 to 72–72.7 Mg/ha in 5- and 10-year-old Eucalyptus globulus or Pinus radiata plantations. Similarly, total carbon stocks in these plantations are 129 and 136.2–142.3 Mg/ha, respectively (Gamarra Ramos 2001, Cuellar Bautista and Salazar Hinostroza 2016, Bernachea Jesus 2019). However, Raboin and Posner (2012) report that a 28-year-old Pinus spp. plantation stores only 35.7 Mg/ha of carbon.
Habitats for biodiversity maintenance
Secondary forest
Post-fire 10-year-old secondary forests in tropical montane cloud forest contain 75–93% of plant species found in unburned areas, and old burned areas (10–28 years) contain 72% more diversity than nearby mature forest (Oliveras et al. 2014). Similarly, secondary forests host 66.7% more dung beetle species than surrounding mature forest (Vélez Quesquén and Saavedra Chávez 2019). In contrast, old secondary forests support 36–105.4% of bird species occurring in mature forests (Colorado Zuluaga and Rodewald 2015, Hosner et al. 2015; Appendix 2).
Agroforestry systems
Shaded coffee agroforestry hosts a greater diversity of bird species (120.3%) than nearby mature forest, but most of the species in agroforestry systems are generalists (Colorado Zuluaga and Rodewald 2015; Appendix 2).
Tree plantations
Compared to mature forests, Eucalyptus and Alnus (alder) plantations host 79–120.8% of shrub and 70.2–84.2% of soil macrofauna species (de Valença et al. 2017). Information for other taxa was not found.
Hydrological services
Water yield and storage: Secondary forests have a slightly higher capacity to buffer peak flows and to store water in the soil than exotic tree plantations (estimated odds ratio: 2.22 vs. 2.37; Bonnesoeur et al. 2019). In contrast, water yield in agroforestry systems (shaded coffee) and forest tree plantations (in former pasture) is 11% and 14% lower than in mature forests, respectively (Quintero et al. 2009). Other studies have shown similar hydrological patterns for tree plantations. Reforestation with exotic trees (Pinus patula, Eucalyptus globulus, Cupressus lusitanica) over 1% of the catchment area resulted in a decrease of 20% and 40% water yield if they replaced grazed or natural grassland, respectively (Bonnesoeur et al. 2019). Similarly, the oldest pine plantations on high Andean grassland retained up to 63% less water than natural grassland soils (Farley et al. 2004) and reduced water yields by ~50% or an average of 242 mm/yr (Buytaert et al. 2007). In contrast, water regulation was only slightly affected in agroforestry and silvopasture (Eucalyptus viminalis and Caesalpinia spinosa) on the western slope of the Andes (Villar Cabeza et al. 2014).
Water infiltration and soil moisture: A 14–20-year-old forestation on degraded soils improves infiltration rates by eight times, but it was three times lower than in mature native forests (Bonnesoeur et al. 2019). Soil moisture content in Polylepis secondary forest (in a former potato field) was slightly lower (45–53%) compared to nearby natural grasslands (50–74%), whereas soil in a 20-year-old pine plantation had significantly lower moisture content (13–22%) than in natural grassland (Harden et al. 2013).
Water flow regulation: Afforestation with pine and Eucalyptus species reduces surface runoff by 9–11% and flow volume by 6–8% in the western Andes caused by higher water use (Krois and Schulte 2013). Water mean daily flows in Eucalyptus afforested areas are 4–10 times lower than in neighboring natural catchments (Ochoa-Tocachi et al. 2016, Ochoa-Tocachi 2019).
Air humidity interception: Relative canopy water intercept is 25% of bulk rainfall in agroforestry, similar to secondary forests and tree plantations, regardless of tree density (Bonnesoeur et al. 2019).
Water quality: Pine plantations change water quality minimally (van Dijk and Keenan 2007).
Similar to secondary forests in the Amazonian region, secondary forests in the Andes host a high diversity of birds, trees, and dung beetles (Box 2, Appendix 2). In tropical montane cloud forests, fires occur with relative frequency and cause severe impacts on biodiversity (Young and León 2007, Oliveras et al. 2014), but rapid and vigorous resprouting in burned areas can occur (Román-Cuesta et al. 2011, Oliveras et al. 2014, 2018), allowing for rapid ex-ante species recovery (Oliveras et al. 2014; Box 2, Fig. 3). In agroforestry systems and tree plantations, managers often employ techniques to control tree density and diversity and to control pests and weeds, which affect other species. In addition, some exotic tree plantations (e.g., Eucalyptus, widely planted in the Andes) release allelopathic compounds that prevent the establishment of native species. These are probably some of the reasons why agroforestry systems and tree plantations contain fewer or mainly generalist insect species (Box 2).
Our literature review reveals that forests in the Andes have either positive or negative effects on hydrological services, depending on the forest type (Box 2). Natural forest regeneration and reforestation with native species improve water supply and regulation services, whereas exotic tree plantations have negative effects on hydrological regulation, especially when they replace natural grasslands (Box 2). These exotic tree plantations reduce stream flow and groundwater recharge because they consume large quantities of water and release it through transpiration. This situation implies a reduction in water availability for downstream users, especially during the dry season (Box 2).
Coastal forest ecosystem services
Although there is little information about dry forests and their ecosystem services in the coastal Peruvian region, it is well known that dry forests are extremely important sources of subsistence and income for local communities (wood, firewood, and food provided by Prosopis species). Coastal dry forests are threatened by deforestation despite their sparse tree presence (< 30% cover in most of the coastal region) and the dominance of algarrobo species (Prosopis pallida; León Caceres 2019). Although dry forests contain lower carbon stocks than Andean and Amazonian forests (Box 3), native vegetation regeneration in this ecosystem, particularly Prosopis species, plays an important role in providing crucial ecosystem services such as water provision services and habitats for biodiversity. For example, dry secondary forests support high endemicity that results in a relatively high diversity of trees and mammals such as insectivorous bats (Box 3). Prosopis trees increase soil humidity by up to 28%, absorbing water from deep soil horizons and capturing a significant volume of water from atmospheric humidity (Box 3). Because rainfall is almost nonexistent on the coast during the long dry season, vegetation provides a significant annual water supply through fog capture and condensation (Whaley et al. 2010). Coastal dry forest plants have evolved adaptations (Box 3) to survive these environmental conditions. Adaptations such as stomatal regulation regulate water consumption and allow the species to endure drought stress (Time et al. 2018); small and lignified leaf branches, or “brachyblats”, capture atmospheric humidity (Whaley et al. 2010); and “inverse hydraulic lift” provides the ability to absorb atmospheric water at night (Bereford-Jones 2004). A recent study shows how important these forests are in reducing the vulnerability of human populations to El Niño, as the loss of dry forest cover magnifies the negative effects of drought and increases local temperatures (Pécastaing and Chávez 2020).
Here, we summarize the main findings for three key ecosystem services provided by secondary forests and post-deforestation forest transitions (agroforestry systems and tree plantations) in the coastal region of Peru. For a complete list of studies reviewed and their carbon stock values and species occurrences see Appendix 1 and 2.
Carbon stocks
Mature forest
Aboveground carbon stocks in coastal dry forest are reported to be on average 27.6 Mg/ha, and total carbon stocks averaged 44.1 Mg/ha (Cuellar Bautista and Salazar Hinostroza 2016).
Secondary forest
The 25-year-old secondary forests store 4.23 Mg/ha of carbon in aboveground biomass (~15% with respect to mature forest) and 46.9 Mg/ha of total carbon stock (similar to mature forest; Chávez Suazo 2018).
Tree plantation
Aboveground carbon stock in Mangifera indica plantations is 8.2 Mg/ha, and total carbon stock is 14.3/ha (Cuellar Bautista and Salazar Hinostroza 2016).
Agroforestry systems
We found no data on carbon stocks for agroforestry systems, suggesting the need for more study in this region.
Aboveground carbon content estimation using LiDAR remote sensing and including all forest types indicates that coastal Peruvian dry forests store on average 1–8 ± 2 Mg/ha of carbon (Asner et al. 2014).
Habitats for biodiversity maintenance
Secondary forests
Peruvian coastal dry secondary forests support 52.2–70% of tree species (Rasal Sánchez et al. 2011, Delgado Paredes et al. 2020) and 166.7% of insectivorous bat species found in nearby mature forest (Ruiz Romero 2015). However, secondary forests in northwestern Peru contain only 22.7% of bird species compared to those in mature forest (León Cáceres 2019; Appendix 2).
Agroforestry systems
Gossypium and Musa agroforestry systems host 79.6% of bird species found in mature dry forest (Chávez-Villavicencio 2013).
Hydrological services
In the arid Peruvian coastal region, Prosopis species are the most representative and common species that play an essential ecological role in water ecosystem services. This function is mediated by following mechanisms:
- Deep dimorphic root systems (up to 60 m) of Prosopis obtain water from deep soil horizons and deposit part of that water along with their dense superficial root network, increasing the upper soil (< 35 cm) moisture by up to 28% (Beresford-Jones 2004, Whaley et al. 2010). This mechanism in which water absorbed by deep roots moves through the roots and is released into the upper soil profile at night (called ‘hydraulic lift’) was demonstrated for several other species growing in arid habitats (Caldwell and Richards 1989, Caldwell et al. 1998, Time et al. 2018). The hydraulically lifted water in arid environments forms a buffer supply to even out water stress during the day for neighboring species and for the lifting plant itself (Beresford-Jones 2004).
- Tiny leaflets in dense clusters of brachyblasts (smallest lignified leaf branches) of Prosopis are adapted to condense atmospheric humidity and precipitate it beneath the canopy (Beresford-Jones 2004, Whaley et al. 2010). For example, a small P. pallida tree (3 m in height with a crown of 4 m) captures up to 9 L of water each night (Whaley et al. 2010). Prosopis also absorbs atmospheric water through its leaves at night and fixes moisture into the soil (by a mechanism called “inverse hydraulic lift”; Beresford-Jones 2004). This mechanism is essential because precipitation is basically nonexistent in many parts of Peruvian coastal regions, and fog can represent up to 50% of water flow (Brandon 2014).
- Prosopis is an efficient user of available water in hyper-arid areas. For example, P. juliflora uses approximately 235 mm/yr of water, which represents only 20% of Eucalyptus camaldulensis water consumption under the same conditions (Mahmood et al. 2001, Beresford-Jones 2004).
Overall, Prosopis generates “islands of humidity” around the plant and can provide important benefits in water provision, both in natural and human-modified forest transitions (Beresford-Jones 2004).
Overall, our synthesis shows that all forest transition types provide valuable services. However, it is important to consider the amount and importance of the ecosystem services provided. Although agroforestry systems and tree plantations provide carbon stocks and wildlife habitat services, they still result in high proportions of ecosystem services losses. Moreover, exotic tree plantations have negative effects on hydrological services, with a consequent reduction in water availability. In contrast, we found that secondary forest is a more effective and immediate forest type to optimize the provision of various ecosystem services, which we explore further below.
Secondary forests as nature-based solutions
Role of secondary forests in mitigating climate change
Compared to the immense effect of the human footprint on the Earth’s ecosystems, climate change has so far played a relatively smaller role. However, the Anthropocene and its human interference has increased the uncertainty of climate patterns (e.g., changing rainfall patterns, extreme temperature fluctuations, higher frequency of extreme events), making it more difficult to mitigate and adapt to climate change, protect biodiversity, and secure human well-being (Malhi et al. 2020). Immediate and ambitious climate change mitigation action is necessary to reduce the severity of the impacts that societies and ecosystems must face. To achieve this goal, nature-based solutions are emerging as integrated approaches that can help to reduce and protect humans from climate change impacts while slowing global warming, supporting biodiversity, and securing ecosystem service provision (Griscom et al. 2017, Chausson et al. 2020, Seddon et al. 2020). Secondary tropical forests play essential roles in nature-based solutions such as natural climate solutions (mitigation potential associated with secondary forests’ higher sequestration rates compared to more mature forests), green infrastructure (secondary forests help prevent erosion and reduce storm impacts), and ecosystem-based adaptation (water storage due to better soil infiltration rates and water storing options such as mosses in Andean forests).
Peru has at least two direct ways to put secondary forests under nature-based solutions for the United Framework Convention on Climate Change (UNFCCC). One would be to place secondary forests’ mitigation potential as a clearly defined component of the country’s intended nationally determined contributions. A second way is under Peru’s commitment to restore 1.2 million ha of degraded land (out of 3.2 million ha total committed under Initiative 20x20). In September 2015, the Peruvian government submitted its first intended nationally determined contributions document to the UNFCCC, committing to an emissions reduction equivalent to 30% below the projected business-as-usual level by 2030 (Gobierno del Perú 2016). Peru has displayed greater ambition in its recently revised resubmission, committing to reduce emissions by 40% in the next decade (Gobierno del Perú 2020). Approximately 70% of the mitigation efforts to achieve this commitment consider changes in land use and improvements to the forestry sector (including afforestation and agricultural efficiency; Gobierno del Perú 2016, Gonzales-Zuñiga 2018, Gallice et al. 2019). However, the Peruvian government is still not adopting this strategy (Climate Action Tracker 2019) and it does not explicitly consider the potential of secondary forests (Gobierno del Perú 2020). As a result, Peru’s current climate policy actions are labelled as “insufficient” and are not consistent with the Paris Agreement’s limit to 1.5°C (Climate Action Tracker 2019). Estimates suggest that improving forest management would lead to achieving 32% of Peru’s current conditional target (Climate Action Tracker 2019). At this time, Peru lacks national policies on secondary forest management. Secondary forests in the Andean and coastal region are even more neglected in terms of study, protection, and sustainable use than Amazonian forests. This lack may be attributed, in part, to the Peruvian Ministry of the Environment, which is still locating and quantifying the carbon storage and mitigation potential of secondary forests across the country. We highlight the urgent need to make a greater effort to accelerate the adoption of policies that promote secondary forest management, standing forest preservation, and inclusion of old-growth and regrowth forests under payment for ecosystem services to stimulate their permanence. One of the common problems with including secondary forests under both nationally determined contributions and Initiative 20x20 commitments is that secondary forests are seen only as carbon. However, these forests offer many more services than only carbon, as shown here and in other studies (Lewis et al. 2019).
Mitigation activities in Peru and elsewhere in Latin America are encouraging low-biodiversity afforestation and reforestation commercial practices with non-native monocultures (Zamorano-Elgueta et al. 2015, Moomaw et al. 2019, Heilmayr et al. 2020, Seddon et al. 2020). This approach can result in a maladaptive practice, especially considering rapid shifts to strategies in which biodiversity-based resilience is key. The carbon-centric approach is also detrimental to the preservation of standing forests, particularly mature old-growth, and places less effort into promoting forest permanence (Lewis et al. 2019).
We highlight that secondary forests are an important cost-effective option for land management that supports multiple environmental commitments that are currently underrepresented in Latin America (Chazdon and Guariguata 2016, 2018). We offer three reasons:
- Trees in secondary forests are already growing and sequestering and storing carbon at high rates (Asner et al. 2010, Chazdon et al. 2016, Poorter et al. 2016), whereas newly planted forests require many years before they sequester carbon dioxide in significant quantities (Box 1–3; Fig. 2A). Furthermore, secondary forests have greater tree species diversity and higher rates of carbon sequestration, whereas plantations are often monocultures and stock and sequester less carbon more slowly than secondary forests, particularly in the Amazonian region. Because carbon dioxide removal rate needs to increase rapidly to meet temperature limitation goals (IPCC 2018), secondary forests are a fast route to sequestering atmospheric carbon. More frequent droughts, insect pests, and diseases can compromise the successful establishment of tree plantations and agroforestry enterprises and their associated benefits.
- Tree plantations and agroforestry systems are mostly planted with commercial purposes and are not designed to result in permanent forest cover. Consequently, these forests could compromise long-term carbon sequestration because trees are harvested after a few decades (Farley et al. 2005, Luzar 2007, Raboin and Posner 2012). In fact, we have not found any study reporting plantations or agroforestry systems older than 30 years. Conflicts between socioeconomic and environmental aspects also influence the persistence of human-managed forests. For example, in a rural community of Cusco (Piñapampa), approximately 80–90 trucks each week deliver Eucalyptus wood to the town for use as fuel (thus releasing the carbon dioxide into the atmosphere), and local inhabitants earn, on average, $60,000–81,000 USD during a typical logging season (Luzar 2007). Considering the costs and benefits, these values are much higher than the potential market based on carbon sequestration (Luzar 2007, Raboin and Posner 2012).
- Vast land areas are frequently needed to optimize afforestation and reforestation benefits, which could potentially result in the destruction of natural habitats and competition with food production, urban space, and other land uses. For example, large areas of natural grasslands in the Peruvian Andes have been forested with exotic Eucalyptus and Pinus species, which are having negative effects on hydrological services and biodiversity conservation (Krois and Schulte 2013, Tovar et al. 2013, Bonnesoeur et al. 2019).
Secondary forests as habitat for biodiversity conservation
Secondary forests are not only effective and cost-effective natural absorbers of carbon, but they also provide essential habitat for many plants and animals, including threatened species that are often absent from human-managed forests. This function is particularly important in ecosystems with high endemicity, such as the Andes and the semideciduous dry forests on the coast. A naturally regenerating forest is undergoing self-organization and an increase in structural complexity and diversity over time, whereas tree plantations and agroforestry systems experience continuous human intervention. Although better than bare land, managed forests can have their conservation goals compromised compared with secondary forests. Biodiversity, in turn, offers fundamental human services, including reduced risk of zoonosis (Gibb et al. 2020). Peru has achieved various objectives of its strategic plan for biodiversity (2011–2020), linked to the Aichi biodiversity conservation targets under the Convention on Biological Diversity: for example, target 11, which states that at least 17% of terrestrial areas must be protected by 2020 (MINAM 2015, Maxwell et al. 2020). The complexity of climate change, however, requires new perspectives on conservation strategies involving not only permanent protected areas but also in combination with corridors and temporary conservation areas to create flexible networks that provide critical transitional areas to biodiversity redistributions (D’Aloia et al. 2019). In current landscapes with mosaics of heterogeneous habitats, secondary forests can be fundamental in connecting conservation areas and may also provide regional habitat heterogeneity and vegetation structural complexity. Along this line, Horváth et al. (2019) highlight that the loss in species richness is exacerbated by habitat loss via connectivity loss, thus reinforcing the importance of habitat connectivity in biodiversity conservation policy. The persistence of secondary forests is fundamental to long-term biodiversity conservation, which in turn helps preserve the multiple services that biodiversity provides to human beings, starting by prevention of pandemics and zoonotic diseases.
Role of secondary forests in hydrological services
Water supply is one of the critical issues facing society in the 21st century. As deforestation and climate change alter weather patterns and their variability, severe water deficits and floods are likely to become more frequent. In this context, vegetation recovery can positively or negatively influence local and regional water supply and regulation. In the case of Peru, secondary forests influence local and regional water supplies, which are particularly important in the Andes. The Andean region is the main water provider (from rainwater and glacier melt) not only to local communities, but also to the dry region along the Pacific coast (Beresford-Jones 2004, Whaley et al. 2010) and the eastern Amazonian region (Clark et al. 2014, Doornbos 2015). Consequently, biodiversity and the livelihoods of millions of people depend directly on water services provided by Andean ecosystems (Doornbos 2015). In addition, water is vital to economically important activities such as agriculture, energy, fishing, and tourism. In Peru, 70% of the water for hydroelectricity generation comes from Andean rivers (Doornbos 2015). Temperatures in the tropical Andes have been increasing, melting glaciers and increasing downstream water supply (Vuille et al. 2008, 2018). Nevertheless, this water increase is temporary and is not sustainable. In contrast, future reductions in water availability for both human consumption and other economically important activities is expected (Vuille et al. 2018). The imminent impacts of climate change on water supply motivated the Peruvian government and nongovernmental organizations to implement a variety of rainwater harvesting techniques, including forestation (Somers et al. 2018, Locatelli et al. 2020). Exotic trees (Eucalyptus and Pinus) planted in large areas of the Andes have negative effects on hydrological services, reducing stream flow and groundwater recharge, and thus reducing water availability for downstream users (Luzar 2007, Ochoa-Tocachi et al. 2016, Ochoa-Tocachi 2019). Furthermore, fast-growing exotic Eucalyptus and Pinus plantations are established in wetlands or on the margins of Andean catchments, resulting in the aforementioned negative effects (Luzar 2007, Ochoa-Tocachi et al. 2016). Thus, redirecting the national restoration strategies is necessary to achieve the desired goals. Exotic tree plantations on natural grasslands must be avoided, and secondary forest persistence and restoration with native species and mature forest conservation should be favored to promote their excellent hydrological control in the Andes (Box 2).
Although we tend to highlight the negative effects of exotic tree species on water yields, if well managed (e.g., implemented in adequate areas such as degraded land far from watersheds), they can play an important economic role and help to control erosion and to stabilize soils against landslides (Guns and Vanacker 2013, Bonnesoeur et al. 2019). In addition, plantations can indirectly enhance some ecosystem services by avoiding further logging of native forests (Bonnesoeur et al. 2019). Finally, it is important to consider that the improvement of hydrological regulation is a slow process that may take decades, making outcomes for society and the environment slow to realize (Bonnesoeur et al. 2019).
CONCLUDING REMARKS
We provide a robust synthesis of published evidence regarding the ecosystem services and disservices of forest transitions to encourage the adoption of policies that promote the sustainable use and conservation of secondary forests in Peruvian landscapes. In this review, we compiled empirical evidence to provide a framework for future research and policy decision-making (Fig. 4A). We found that forest transitions provide significant services in offsetting carbon emissions, regulating hydrological services, and providing habitats for biodiversity. However, the amount and importance of ecosystem services vary depending on the forest transition type. Secondary forests offer multiple cost-effective services from climate change mitigation and water regulation to species conservation through the maintenance of landscape connectivity, regional heterogeneity, and vegetation structural complexity.
Our findings highlight the missed opportunity of relying on secondary forests to implement more ambitious and integrated environmental initiatives such as the Convention on Biodiversity-Aichi biodiversity targets, UNFCCC-NDC-REDD+, and Initiative 20x20 for land restoration. Current commitments include high-cost strategies, including potential disservices (e.g., two million commercial tree plantations under Initiative 20x20). In contrast, secondary forests are a low-cost, near-term, highly effective strategy for mitigating the crises in climate and biodiversity and ultimately providing vital ecosystem services to society, such as water provision. We urge considering both ecosystem services and disservices, as well as valuing the relative importance of different land options for each specific region (Fig. 4) to avoid trade-offs in which the achievement of one goal is detrimental to another. Policy-makers and practitioners can use our synthesis to support decisions on which transitions types are best to achieve specific end goals at local and regional scales in Peru. Our synthesis can serve to enhance the visibility and underscore the usefulness of strategies to preserve secondary forests, as well as mature standing forests. Finally, it is also important to consider that the persistence of secondary forests is closely linked to costs and benefits in which the interests of local people in conserving the forest areas can be decisive (Fig. 4A).
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.AUTHOR CONTRIBUTIONS
RT, NS, and RMRC conceived the idea; RT performed the review; RT, NS, EGC, TEBE, and RMRC wrote the draft; and all authors discussed and contributed to the final version of the manuscript.
ACKNOWLEDGMENTS
This work was supported by CONCYTEC (Peru), World Bank grant (contract 011-2019-FONDECYT-BM-INC-INV). We thank Dr. José C. Silva and three anonymous reviewers for useful suggestions made on earlier drafts of the manuscript.
DATA AVAILABILITY
All data used in this manuscript are available in Appendixes 1 and 2.
LITERATURE CITED
Aerts, R., S. Spranghers, and Ç. H. Şekercioğlu. 2017. Conservation of ecosystem services does not secure the conservation of birds in a Peruvian shade coffee landscape. Bird Conservation International 27(1):71-82. https://doi.org/10.1017/S0959270916000149
Aide, T. M., H. R. Grau, J. Graesser, M. J. Andrade-Nuñez, E. Aráoz, A. P. Barros, M. Campos-Cerqueira, E. Chacon-Moreno, F. Cuesta, R. Espinoza, M. Peralvo, M. H. Polk, X. Rueda, A. Sanchez, K. R. Young, L. Zarbá, and K. S. Zimmerer. 2019. Woody vegetation dynamics in the tropical and subtropical Andes from 2001 to 2014: satellite image interpretation and expert validation. Global Change Biology 25(6):2112-2126. https://doi.org/10.1111/gcb.14618
Alegre, J., L. Arévalo, and A. Ricse. 2003. Reservas de carbono según el uso de la tierra en dos sitios de la Amazonia Peruana. In M. D. Sánchez an M. R. Méndez, editors. Agroforestería para la producción animal en América Latina - II - memorias de la segunda conferencia electrónica (Agosto de 2000–Marzo de 2001). Food and Agriculture Organization, Rome, Italy. https://www.fao.org/3/Y4435S/Y4435S00.htm
Álvarez-Berríos, N., J. L’Roe, and L. Naughton-Treves. 2021. Does formalizing artisanal gold mining mitigate environmental impacts? Deforestation evidence from the Peruvian Amazon. Environmental Research Letters 16(6):064052. https://doi.org/10.1088/1748-9326/abede9
Aragón, S., N. Salinas, A. Nina-Quispe, V. H. Qquellon, G. R. Paucar, W. Huaman, P. C. Porroa, J. C. Olarte, R. Cruz, J. G. Muñiz, C. S. Yupayccana, T. E. B. Espinoza, R. Tito, E. G. Cosio, and R. M. Roman-Cuesta. 2021. Aboveground biomass in secondary montane forests in Peru: slow carbon recovery in agroforestry legacies. Global Ecology and Conservation 28:e01696. https://doi.org/10.1016/j.gecco.2021.e01696
Arévalo-Gardini, E., M. Canto, J. Alegre, O. Loli, A. Julca, and V. Baligar. 2015. Changes in soil physical and chemical properties in long term improved natural and traditional agroforestry management systems of cacao genotypes in Peruvian Amazon. Plos One 10(7):e0132147. https://doi.org/10.1371/journal.pone.0132147
Asner, G. P., D. E. Knapp, R. E. Martin, R. Tupayachi, C. B. Anderson, J. Mascaro, F. Sinca, K. D. Chadwick, S. Sousan, M. Higgins, W. Farfan, M. R. Silman, W. A. Llactayo León, and A. F. Neyra Palomino. 2014. La geografía del carbono en alta resolución del Perú. Carnegie Institution for Science, Washington, D.C., USA. https://geoservidorperu.minam.gob.pe/geoservidor/Archivos/Documentos/CarnegiePeruCarbonReport-Spanish.pdf
Asner, G. P., G. V. N. Powell, J. Mascaro, D. E. Knapp, J. K. Clark, J. Jacobson, T. Kennedy-Bowdoin, A. Balaji, G. Paez-Acosta, E. Victoria, L. Secada, M. Valqui, and R. F. Hughes. 2010. High-resolution forest carbon stocks and emissions in the Amazon. Proceedings of the National Academy of Sciences 107(38):16738-16742. https://doi.org/10.1073/pnas.1004875107
Baltazar Peña, A. L. 2011. Carbono aéreo almacenado en una plantación de bolaina blanca (Guazuma crinita Martius) de un año de edad, en el centro poblado Nuevo Edén, alto Madre de Dios. Thesis. Universidad Nacional Agraria De La Selva, Tingo María, Perú. http://repositorio.unas.edu.pe/handle/UNAS/563
Barbarán, G. 1998. Determinación de biomasa y carbono en los principales sistemas de uso del suelo en la zona de Campo Verde. Thesis. Universidad Nacional de Ucayali, Pucallpa, Perú.
Barbier, E. B., J. C. Burgess, and A. Grainger. 2010. The forest transition: towards a more comprehensive theoretical framework. Land Use Policy 27(2):98-107. https://doi.org/10.1016/j.landusepol.2009.02.001
Barlow, J., T. A. Gardner, I. S. Araujo, T. C. Ávila-Pires, A. B. Bonaldo, J. E. Costa, M. C. Esposito, L. V. Ferreira, J. Hawes, M. I. M. Hernandez, M. S. Hoogmoed, R. N. Leite, N. F. Lo-Man-Hung, J. R. Malcolm, M. B. Martins, L. A. M. Mestre, R. Miranda-Santos, A. L. Nunes-Gutjahr, W. L. Overal, L. Parry, S. L. Peters, M. A. Ribeiro-Junior, M. N. F. da Silva, C. da Silva Motta, and C. A. Peres. 2007. Quantifying the biodiversity value of tropical primary, secondary, and plantation forests. Proceedings of the National Academy of Sciences 104(47):18555-18560. https://doi.org/10.1073/pnas.0703333104
Bennett-Curry, A., Y. Malhi, and M. Menton. 2013. Leakage effects in natural resource supply chains: a case study from the Peruvian commercial charcoal market. International Journal of Sustainable Development and World Ecology 20(4):336-348. https://doi.org/10.1080/13504509.2013.804892
Beresford-Jones, D. G. 2004. Pre-Hispanic Prosopis-human relationships on the south coast of Peru: riparian forests in the context of environmental and cultural trajectories of the lower Ica valley. Dissertation. University of Cambridge, Cambridge, UK.
Bernachea Jesus, N. 2019. Valoración económica y secuestro de CO2 en bosques plantados de eucalipto (Eucalyptus globulus labil) y pino (Pinus radiata) de 11 años de edad en Cochatama, distrito Huacar, provincia Ambo departamento Huánuco – Setiembre, 2018 – Febrero, 2019. Thesis. Universidad de Huánuco, Huánuco, Perú. http://repositorio.udh.edu.pe/123456789/1776
Bonnesoeur, V., B. Locatelli, M. R. Guariguata, B. F. Ochoa-Tocachi, V. Vanacker, Z. Mao, A. Stokes, and S.-L. Mathez-Stiefel. 2019. Impacts of forests and forestation on hydrological services in the Andes: a systematic review. Forest Ecology and Management 433:569-584. https://doi.org/10.1016/j.foreco.2018.11.033
Bourrel, L., P. Rau, B. Dewitte, D. Labat, W. Lavado, A. Coutaud, A. Vera, A. Alvarado, and J. Ordoñez. 2015. Low-frequency modulation and trend of the relationship between ENSO and precipitation along the northern to centre Peruvian Pacific coast. Hydrological Processes 29(6):1252-1266. https://doi.org/10.1002/hyp.10247
Brandon, K. 2014. Ecosystem services from tropical forests: review of current science. Center for Global Development Working Paper 380. Center for Global Development, Washington, D.C., USA. https://doi.org/10.2139/ssrn.2622749
Bruijnzeel, L. A. 2004. Hydrological functions of tropical forests: not seeing the soil for the trees? Agriculture, Ecosystems and Environment 104(1):185-228. https://doi.org/10.1016/j.agee.2004.01.015
Bullock, E. L., C. E. Woodcock, and P. Olofsson. 2020. Monitoring tropical forest degradation using spectral unmixing and Landsat time series analysis. Remote Sensing of Environment 238:110968. https://doi.org/10.1016/j.rse.2018.11.011
Butler, R. A., and W. F. Laurance. 2009. Is oil palm the next emerging threat to the Amazon? Tropical Conservation Science 2(1):1-10. https://doi.org/10.1177%2F194008290900200102
Buytaert, W., V. Iñiguez, and B. De Bièvre. 2007. The effects of afforestation and cultivation on water yield in the Andean páramo. Forest Ecology and Management 251(1-2):22-30. https://doi.org/10.1016/j.foreco.2007.06.035
Caballero Espejo, J., M. Messinger, F. Román-Dañobeytia, C. Ascorra, L. E. Fernandez, and M. Silman. 2018. Deforestation and forest degradation due to gold mining in the Peruvian Amazon: a 34-year perspective. Remote Sensing 10(12):1903. https://doi.org/10.3390/rs10121903
Caldwell, M. M., T. E. Dawson, and J. H. Richards. 1998. Hydraulic lift: consequences of water efflux from the roots of plants. Oecologia 113(2):151-161. https://doi.org/10.1007/s004420050363
Caldwell, M. M., and J. H. Richards. 1989. Hydraulic lift: water efflux from upper roots improves effectiveness of water uptake by deep roots. Oecologia 79(1):1-5. https://doi.org/10.1007/BF00378231
Cerrón, J., J. del Castillo, S. L. Mathez-Stiefel, and E. Thomas. 2017. Lecciones aprendidas de experiencias de restauración en el Perú. Biodiversity International, Lima, Peru. https://www.bioversityinternational.org/e-library/publications/detail/lecciones-aprendidas-de-experiencias-de-restauracion-en-el-peru/
Cerrón, J., T. Fremout, R. Atkinson, E. Thomas, and J. Cornelius. 2019. Experiencias de restauración y fuentes semilleras en el bosque seco tropical del norte del Perú: estado actual y lecciones aprendidas. Biodiversity International, Lima, Peru. https://www.bioversityinternational.org/e-library/publications/detail/experiencias-de-restauracion-y-fuentes-semilleras-en-el-bosque-seco-tropical-del-norte-del-peru-est/
Chausson, A., B. Turner, D. Seddon, N. Chabaneix, C. A. J. Girardin, V. Kapos, I. Key, D. Roe, A. Smith, S. Woroniecki, and N. Seddon. 2020. Mapping the effectiveness of nature-based solutions for climate change adaptation. Global Change Biology 26(11):6134-6155. https://doi.org/10.1111/gcb.15310
Chávez Suazo, J. P. 2018. Captura y almacenamiento de carbono de los bosque estacionalmente secos de la costa norte (Lambayeque), como mitigación frente al cambio climático. Thesis. Universidad Nacional Agraria la Molina, Lima, Peru. https://hdl.handle.net/20.500.12996/3346
Chávez-Villavicencio, C. 2013. Las aves de la ciudad de Piura, Perú y sus alrededores: ocho años mirando al cielo. Biologist 11(2):193-204. https://doi.org/10.24039/rtb2013112398
Chazdon, R. L., E. N. Broadbent, D. M. A. Rozendaal, F. Bongers, A. M. A. Zambrano, T. M. Aide, P. Balvanera, J. M. Becknell, V. Boukili, P. H. S. Brancalion, D. Craven, J. S. Almeida-Cortez, G. A. L. Cabral, B. de Jong, J. S. Denslow, D. H. Dent, S. J. DeWalt, J. M. Dupuy, S. M. Durán, M. M. Espírito-Santo, M. C. Fandino, R. G. César, J. S. Hall, J. L. Hernández-Stefanoni, C. C. Jakovac, A. B. Junqueira, D. Kennard, S. G. Letcher, M. Lohbeck, M. Martínez-Ramos, P. Massoca, J. A. Meave, R. Mesquita, F. Mora, R. Muñoz, R. Muscarella, Y. R. F. Nunes, S. Ochoa-Gaona, E. Orihuela-Belmonte, M. Peña-Claros, E. A. Pérez-García, D. Piotto, J. S. Powers, J. Rodríguez-Velazquez, I. E. Romero-Pérez, J. Ruíz, J. G. Saldarriaga, A. Sanchez-Azofeifa, N. B. Schwartz, M. K. Steininger, N. G. Swenson, M. Uriarte, M. van Breugel, H. van der Wal, M. D. M. Veloso, H. Vester, I. C. G. Vieira, T. V. Bentos, G. B. Williamson, and L. Poorter. 2016. Carbon sequestration potential of second-growth forest regeneration in the Latin American tropics. Science Advances 2(5):e1501639. https://doi.org/10.1126/sciadv.1501639
Chazdon, R. L., and M. R. Guariguata. 2016. Natural regeneration as a tool for large-scale forest restoration in the tropics: prospects and challenges. Biotropica 48(6):716-730. https://doi.org/10.1111/btp.12381
Chazdon, R. L., and M. R. Guariguata. 2018. Decision support tools for forest landscape restoration: current status and future outlook. Occasional Paper 183. Center for International Forestry Research, Bogor, Indonesia. https://doi.org/10.17528/cifor/006792
Chazdon, R. L., D. Lindenmayer, M. R. Guariguata, R. Crouzeilles, J. M. Rey Benayas, and E. Lazos Chavero. 2020. Fostering natural forest regeneration on former agricultural land through economic and policy interventions. Environmental Research Letters 15(4):043002. https://doi.org/10.1088/1748-9326/ab79e6
Chuquizuta, P. D., G. F. Ruiz, C. T. Salas, and L. A. López. 2016. Carbono almacenado en cinco sistemas de uso de tierra, en la región San Martín Perú. Rinderesu 1(2):57-67. http://rinderesu.com/index.php/rinderesu/article/view/22
Clark, K. E., M. A. Torres, A. J. West, R. G. Hilton, M. New, A. B. Horwath, J. B. Fisher, J. M. Rapp, A. Robles Caceres, and Y. Malhi. 2014. The hydrological regime of a forested tropical Andean catchment. Hydrology and Earth System Sciences 18(12):5377-5397. https://doi.org/10.5194/hess-18-5377-2014
Climate Action Tracker. 2019. Peru nationally determined contribution (NDC) assessment. Climate Action Network, Bonn, Germany. https://climateactiontracker.org/countries/peru/2019-09-19/
Colorado Zuluaga, G. J., and A. D. Rodewald. 2015. Response of mixed-species flocks to habitat alteration and deforestation in the Andes. Biological Conservation 188:72-81. https://doi.org/10.1016/j.biocon.2015.02.008
Concha, J. Y., J. C. Alegre, and V. Pocomucha. 2007. Determinación de las reservas de carbono en la biomasa aérea de sistemas agroforestales de Theobroma cacao L. en el departamento de San Martin, Perú. Ecología Aplicada 6(1-2):75-82. http://www.scielo.org.pe/scielo.php?pid=S1726-22162007000100009&script=sci_abstract
Csillik, O., and G. P. Asner. 2020. Aboveground carbon emissions from gold mining in the Peruvian Amazon. Environmental Research Letters 15(1):014006. https://doi.org/10.1088/1748-9326/ab639c
Cuellar Bautista, J. E., and E. J. Salazar Hinostroza. 2016. Dinámica del carbono almacenado en los diferentes sistemas de uso de la tierra del Perú, base para una estrategia de mitigación ante el cambio climático. Instituto Nacional de Innovación Agraria, Lima, Peru. http://repositorio.inia.gob.pe/handle/20.500.12955/363
Culot, L., D. J. Mann, F. J. J. Muñoz Lazo, M.-C. Huynen, and E. W. Heymann. 2011. Tamarins and dung beetles: an efficient diplochorous dispersal system in the Peruvian Amazonia. Biotropica 43(1):84-92. https://doi.org/10.1111/j.1744-7429.2010.00655.x
D’Almeida, C., C. J. Vörösmarty, G. C. Hurtt, J. A. Marengo, S. L. Dingman, and B. D. Keim. 2007. The effects of deforestation on the hydrological cycle in Amazonia: a review on scale and resolution. International Journal of Climatology 27(5):633-647. https://doi.org/10.1002/joc.1475
D’Aloia, C. C., I. Naujokaitis-Lewis, C. Blackford, C. Chu, J. M. R. Curtis, E. Darling, F. Guichard, S. J. Leroux, A. C. Martensen, B. Rayfield, J. M. Sunday, A. Xuereb, and M.-J. Fortin. 2019. Coupled networks of permanent protected areas and dynamic conservation areas for biodiversity conservation under climate change. Frontiers in Ecology and Evolution 7:27. https://doi.org/10.3389/fevo.2019.00027
de Valença, A. W., S. J. Vanek, K. Meza, R. Ccanto, E. Olivera, M. Scurrah, E. A. Lantinga, and S. J. Fonte. 2017. Land use as a driver of soil fertility and biodiversity across an agricultural landscape in the central Peruvian Andes. Ecological Applications 27(4):1138-1154. https://doi.org/10.1002/eap.1508
Delgado Paredes, G. E., C. Vásquez-Díaz, F. Tesén-Núñez, B. Esquerre-Ibañez, F. Z. Da-Silva, and C. Rojas-Idrogo. 2020. Vegetación arbórea del Cerro Tres Puntas de Pilasca, (Salas-Motupe), Lambayeque, Perú. Revista Mexicana de Ciencias Forestales 11(58):154-181. https://doi.org/10.29298/rmcf.v11i58.549
Doornbos, B. 2015. El valor de los bosques andinos en asegurar agua y suelo en un contexto de creciente riesgo climático: ¿(re)conocemos lo imperdible? Opinion Article 3. Programa Bosques Andinos, Lima, Peru. https://www.bosquesandinos.org/wp-content/uploads/2015/12/061115_articulo_n3.pdf
Duran-Bautista, E. H., I. Armbrecht, A. N. Serrão Acioli, J. C. Suárez, M. Romero, M. Quintero, and P. Lavelle. 2020. Termites as indicators of soil ecosystem services in transformed amazon landscapes. Ecological Indicators 117:106550. https://doi.org/10.1016/j.ecolind.2020.106550
Ehrenbergerová, L., E. Cienciala, A. Kučera, L. Guy, and H. Habrová. 2016. Carbon stock in agroforestry coffee plantations with different shade trees in Villa Rica, Peru. Agroforestry Systems 90(3):433-445. https://doi.org/10.1007/s10457-015-9865-z
Ellison, D., M. N. Futter, and K. Bishop. 2012. On the forest cover–water yield debate: from demand- to supply-side thinking. Global Change Biology 18(3):806-820. https://doi.org/10.1111/j.1365-2486.2011.02589.x
Escobal, J., and M. Torero. 2003. Adverse geography and differences in welfare in Peru. Pages 77-121 in R. Kanbur and A. J. Venables, editors. Spatial inequality and development. Oxford University Press, Oxford, UK. https://doi.org/10.1093/0199278636.003.0005
Escobedo Grandez, A. 2021. Agronegocios y crisis climática en el Perú: monocultivos, deforestación y emisiones de carbono en la amazonía peruana. Oxfam América, Lima, Peru. https://peru.oxfam.org/latest/policy-paper/agronegocios-y-crisis-climatica-en-peru
Farley, K. A., E. G. Jobbágy, and R. B. Jackson. 2005. Effects of afforestation on water yield: a global synthesis with implications for policy. Global Change Biology 11(10):1565-1576. https://doi.org/10.1111/j.1365-2486.2005.01011.x
Farley, K. A., E. F. Kelly, and R. G. M. Hofstede. 2004. Soil organic carbon and water retention after conversion of grasslands to pine plantations in the Ecuadorian Andes. Ecosystems 7(7):729-739. https://doi.org/10.1007/s10021-004-0047-5
Fitts, L. A., Z. A. Cruz-Burga, and M. La Torre-Cuadros. 2020. Wild rubber extraction in the Peruvian Amazon: local perception and socioeconomic indicators as tools for decision-making. Ethnobiology and Conservation 9:24. https://doi.org/10.15451/ec2020-06-9.24-1-26
Food and Agriculture Organization (FAO). 2016. Bosques y el cambio climático: documento de trabajo 14. FAO, Rome, Italy. https://www.fao.org/documents/card/en/c/50d7476c-8ac1-40d8-a704-db7ed0281662/
Food and Agriculture Organization (FAO). 2020. Global forest resources assessment 2020: main report. FAO, Rome, Italy. https://www.fao.org/documents/card/en/c/ca9825en
Gallice, G. R., G. Larrea-Gallegos, and I. Vázquez-Rowe. 2019. The threat of road expansion in the Peruvian Amazon. Oryx 53(2):284-292. https://doi.org/10.1017/S0030605317000412
Gamarra Ramos, J. 2001. Estimación del contenido de carbono en plantaciones de Eucalyptus globulus Labill, en Junin, Perú. In Simposio internacional medición y monitoreo de la captura de carbono en ecosistemas forestales. Universidad Autónoma de Chihuahua, Chihuahua, Mexico.
Garcia, C. A., S. Savilaakso, R. W. Verburg, V. Gutierrez, S. J. Wilson, C. B. Krug, M. Sassen, B. E. Robinson, H. Moersberger, B. Naimi, J. M. Rhemtulla, H. Dessard, V. Gond, C. Vermeulen, F. Trolliet, J. Oszwald, F. Quétier, S. A. Pietsch, J.-F. Bastin, A. Dray, M. B. Araújo, J. Ghazoul, and P. O. Waeber. 2020. The global forest transition as a human affair. One Earth 2(5):417-428. https://doi.org/10.1016/j.oneear.2020.05.002
Gavin, M. C. 2004. Changes in forest use value through ecological succession and their implications for land management in the Peruvian Amazon. Conservation Biology 18(6):1562-1570. https://doi.org/10.1111/j.1523-1739.2004.00241.x
Gavin, M. C. 2007. Foraging in the fallows: hunting patterns across a successional continuum in the Peruvian Amazon. Biological Conservation 134(1):64-72. https://doi.org/10.1016/j.biocon.2006.07.011
Giam, X. 2017. Global biodiversity loss from tropical deforestation. Proceedings of the National Academy of Sciences 114(23):5775-5777. https://doi.org/10.1073/pnas.1706264114
Gibb, R., D. W. Redding, K. Q. Chin, C. A. Donnelly, T. M. Blackburn, T. Newbold, and K. E. Jones. 2020. Zoonotic host diversity increases in human-dominated ecosystems. Nature 584(7821):398-402. https://doi.org/10.1038/s41586-020-2562-8
Gobierno del Perú. 2016. La contribución nacional del Perú - iNDC: agenda para un desarrollo climáticamente responsable. Government of Peru, Lima, Peru. https://www.minam.gob.pe/cambioclimatico/wp-content/uploads/sites/11/2015/12/LA-CONTRIBUCI%c3%93N-NACIONAL-DEL-PER%c3%9a1.pdf
Gobierno del Perú. 2020. Contribuciones determinadas a nivel nacional del Perú: reporte de actualización periodo 2021–2030. Government of Peru, Lima, Peru. https://cdn.www.gob.pe/uploads/document/file/1675213/Reporte%20de%20
Actualizaci%C3%B3n%20de%20las%20NDC%20del%20Per%C3%BA%20al%202030.pdf
Gonzalez, P., B. Kroll, and C. R. Vargas. 2014. Tropical rainforest biodiversity and aboveground carbon changes and uncertainties in the Selva Central, Peru. Forest Ecology and Management 312:78-91. https://doi.org/10.1016/j.foreco.2013.10.019
Gonzales-Zuñiga, S. 2018. Implementation of nationally determined contributions: Peru country report. Umweltbundesamt, Dessau-Roßlau, Germany. https://www.umweltbundesamt.de/sites/default/files/medien/1410/publikationen/2018-11-30_climate-change_31-2018_country-report-peru.pdf
Griscom, B. W., J. Adams, P. W. Ellis, R. A. Houghton, G. Lomax, D. A. Miteva, W. H. Schlesinger, D. Shoch, J. V. Siikamäki, P. Smith, P. Woodbury, C. Zganjar, A. Blackman, J. Campari, R. T. Conant, C. Delgado, P. Elias, T. Gopalakrishna, M. R. Hamsik, M. Herrero, J. Kiesecker, E. Landis, L. Laestadius, S. M. Leavitt, S. Minnemeyer, S. Polasky, P. Potapov, F. E. Putz, J. Sanderman, M. Silvius, E. Wollenberg, and J. Fargione. 2017. Natural climate solutions. Proceedings of the National Academy of Sciences 114(44):11645-11650. https://doi.org/10.1073/pnas.1710465114
Guns, M., and V. Vanacker. 2013. Forest cover change trajectories and their impact on landslide occurrence in the tropical Andes. Environmental Earth Sciences 70(7):2941-2952. https://doi.org/10.1007/s12665-013-2352-9
Gurmendi Porras, C. G., and W. B. Orihuela Izaguirre. 2019. Valoración económica de la reserva de carbono de Escallonia resinosa (Ruiz & Pav.) Pers. en San Pedro de Saños y Pucará. Universidad Nacional del Centro del Perú, Huancayo, Peru. http://hdl.handle.net/20.500.12894/5320
Hansen, M. C., P. V. Potapov, R. Moore, M. Hancher, S. A. Turubanova, A. Tyukavina, D. Thau, S. V. Stehman, S. J. Goetz, T. R. Loveland, A. Kommareddy, A. Egorov, L. Chini, C. O. Justice, and J. R. G. Townshend. 2013. High-resolution global maps of 21st-century forest cover change. Science 342(6160):850-853. https://doi.org/10.1126/science.1244693
Harden, C. P., J. Hartsig, K. A. Farley, J. Lee, and L. L. Bremer. 2013. Effects of land-use change on water in Andean páramo grassland soils. Annals of the Association of American Geographers 103(2):375-384. https://doi.org/10.1080/00045608.2013.754655
Heilmayr, R., C. Echeverría, and E. F. Lambin. 2020. Impacts of Chilean forest subsidies on forest cover, carbon and biodiversity. Nature Sustainability 3(9):701-709. https://doi.org/10.1038/s41893-020-0547-0
Holl, K. D., and P. H. S. Brancalion. 2020. Tree planting is not a simple solution. Science 368(6491):580-581. https://doi.org/10.1126/science.aba8232
Hölscher, D., J. Mackensen, and J.-M. Roberts. 2005. Forest recovery in the humid tropics: changes in vegetation structure, nutrient pools and the hydrological cycle. Pages 598-621 in M. Bonell and L. A. Bruijnzeel, editors. Forests, water and people in the humid tropics: past, present and future hydrological research for integrated land and water management. Cambridge University Press, Cambridge, UK. https://doi.org/10.1017/CBO9780511535666.031
Horgan, F. G. 2009. Invasion and retreat: shifting assemblages of dung beetles amidst changing agricultural landscapes in central Peru. Biodiversity and Conservation 18(13):3519. https://doi.org/10.1007/s10531-009-9658-7
Horváth, Z., R. Ptacnik, C. F. Vad, and J. M. Chase. 2019. Habitat loss over six decades accelerates regional and local biodiversity loss via changing landscape connectance. Ecology Letters 22(6):1019-1027. https://doi.org/10.1111/ele.13260
Hosner, P. A., M. J. Andersen, M. B. Robbins, A. Urbay-Tello, L. Cueto-Aparicio, K. Verde-Guerra, L. A. Sánchez-González, A. G. Navarro-Sigüenza, R. L. Boyd, J. Núñez, J. Tiravanti, M. Combe, H. L. Owens, and A. Townsend Peterson. 2015. Avifaunal surveys of the upper Apurímac River valley, Ayacucho and Cuzco Departments, Peru: new distributional records and biogeographic, taxonomic, and conservation implications. Wilson Journal of Ornithology 127(4):563-581. https://doi.org/10.1676/14-178.1
International Centre for Research in Agroforestry (ICRAF). 1998. Respuesta a nuevas demandas tecnológicas, fortalecimiento de la investigación en agroindustria y el manejo de recursos naturales. Reporte final enero 1996 a junio 1998. Banco Interamericano de Desarrollo and International Centre for Research in Agroforestry, Lima, Peru.
Intergovernmental Panel on Climate Change (IPCC). 2018. Sumary for policymakers. Pages 3-24 in V. Masson-Delmotte, P. Zhai, H. O. Pörtner, D. Roberts, J. Skea, P. R. Shukla, A. Pirani, W. Moufouma-Okia, C. Péan, R. Pidcock, S. Connors, J. B. R. Matthews, Y. Chen, X. Zhou, M. I. Gomis, E. Lonnoy, T. Maycock, M. Tignor, and T. Waterfield, editors. Global warming of 1.5°C. An IPCC special report on the impacts of global warming of 1.5°C above pre-industrial levels and related global greenhouse gas emission pathways, in the context of strengthening the global response to the threat of climate change, sustainable development, and efforts to eradicate poverty. Cambridge University Press, Cambridge, UK. https://doi.org/10.1017/9781009157940.001
Jezeer, R. E., M. J. Santos, P. A. Verweij, R. G. A. Boot, and Y. Clough. 2019. Benefits for multiple ecosystem services in Peruvian coffee agroforestry systems without reducing yield. Ecosystem Services 40:101033. https://doi.org/10.1016/j.ecoser.2019.101033
Jones, I. L., S. J. DeWalt, O. R. Lopez, L. Bunnefeld, Z. Pattison, and D. H. Dent. 2019. Above- and belowground carbon stocks are decoupled in secondary tropical forests and are positively related to forest age and soil nutrients respectively. Science of the Total Environment 697:133987. https://doi.org/10.1016/j.scitotenv.2019.133987
Krois, J., and A. Schulte. 2013. Modeling the hydrological response of soil and water conservation measures in the Ronquillo watershed in the northern Andes of Peru. Pages 147-184 in Water and environmental dynamics: 6th international conference on water resources and environment research: proceedings. World Meteorological Organization, Koblenz, Germany. https://doi.bafg.de/BfG/2014/ICWRER_2013.pdf
Lapeyre, T., J. Alegre, and L. Arévalo. 2004. Determinación de las reservas de carbono de la biomasa aérea, en diferentes sistemas de uso de la tierra en San Martín, Perú. Ecología Aplicada 3(1-2):35-44. https://doi.org/10.21704/rea.v3i1-2.269
León Cáceres, F. J. 2019. Efectos ambientales por deforestación del bosque seco “El Algarrobal” de Conache -Laredo, 2016. Thesis. Universidad Nacional de Trujillo, Trujillo, Peru. http://dspace.unitru.edu.pe/handle/UNITRU/12919
Lewis, S. L., C. E. Wheeler, E. T. A. Mitchard, and A. Koch. 2019. Restoring natural forests is the best way to remove atmospheric carbon. Nature 568(7750):25-28. https://doi.org/10.1038/d41586-019-01026-8
Locatelli, B., J.-M. Homberger, B. F. Ochoa-Tocachi, V. Bonnesoeur, F. Román, F. Drenkhan, and W. Buytaert. 2020. Impactos de las zanjas de infiltración en el agua y los suelos: ¿Qué sabemos? Forest Trends, Lima, Peru. https://www.forest-trends.org/wp-content/uploads/2020/05/Impactos-de-las-zanjas-de-infiltraci%C3%B3n-en-el-agua-y-los-suelos.pdf
Lojka, B., J. Krausová, Š. Kubík, and Z. Polesný. 2011. Assessment of insect biological diversity in various land use systems in the Peruvian Amazon. Pages 103-121 in N. Rojas and R. Prieto, editors. Amazon basin: plant life, wildlife and environment. Nova Science Publishers, New York, New York, USA.
Luzar, J. 2007. The political-ecology of a “forest transition”: Eucalyptus forestry in the southern Peruvian Andes. Ethnobotany Research and Applications 5:85-93. https://doi.org/10.17348/era.5.0.85-93
MacDonald, H., and D. McKenney. 2020. Envisioning a global forest transition: status, role, and implications. Land Use Policy 99:104808. https://doi.org/10.1016/j.landusepol.2020.104808
Mahmood, K., J. Morris, J. Collopy, and P. Slavich. 2001. Groundwater uptake and sustainability of farm plantations on saline sites in Punjab province, Pakistan. Agricultural Water Management 48(1):1-20. https://doi.org/10.1016/S0378-3774(00)00114-1
Málaga, N., K. Hergoualc’h, G. Kapp, and C. Martius. 2021. Variation in vegetation and ecosystem carbon stock due to the conversion of disturbed forest to oil palm plantation in Peruvian Amazonia. Ecosystems 24(2):351-369. https://doi.org/10.1007/s10021-020-00521-8
Malhi, Y., J. Franklin, N. Seddon, M. Solan, M. G. Turner, C. B. Field, and N. Knowlton. 2020. Climate change and ecosystems: threats, opportunities and solutions. Philosophical Transactions of the Royal Society B 375:20190104. https://doi.org/10.1098/rstb.2019.0104
Manoli, G., A. Meijide, N. Huth, A. Knohl, Y. Kosugi, P. Burlando, J. Ghazoul, and S. Fatichi. 2018. Ecohydrological changes after tropical forest conversion to oil palm. Environmental Research Letters 13(6):064035. https://doi.org/10.1088/1748-9326/aac54e
Marengo, J. A. 2006. On the hydrological cycle of the Amazon basin: a historical review and current state-of-the-art. Revista Brasileira de Meteorologia 21(3):1-19.
Martel, C., and L. Cairampoma. 2012. Cuantificación del carbono almacenado en formaciones vegetales amazónicas en “cicra”, Madre de Dios (Perú). Ecología Aplicada 11(1-2):59-65. https://doi.org/10.21704/rea.v11i1-2.426
Maxwell, S. L., V. Cazalis, N. Dudley, M. Hoffmann, A. S. L. Rodrigues, S. Stolton, P. Visconti, S. Woodley, N. Kingston, E. Lewis, M. Maron, B. B. N. Strassburg, A. Wenger, H. D. Jonas, O. Venter, and J. E. M. Watson. 2020. Area-based conservation in the twenty-first century. Nature 586(7828):217-227. https://doi.org/10.1038/s41586-020-2773-z
McGee, K. M., T. M. Porter, M. Wright, and M. Hajibabaei. 2020. Drivers of tropical soil invertebrate community composition and richness across tropical secondary forests using DNA metasystematics. Scientific Reports 10:18429. https://doi.org/10.1038/s41598-020-75452-4
Ministerio del Ambiente (MINAM). 2014. Estrategia nacional de diversidad biológica al 2021: plan de acción 2014–2018. Ministerio del Ambiente, Lima, Peru. https://sinia.minam.gob.pe/documentos/estrategia-nacional-diversidad-biologica-2021-plan-accion-2014-2018
Ministerio del Ambiente (MINAM). 2015. Mapa nacional de cobertura vegetal: memoria descriptiva. Ministerio del Ambiente, Lima, Peru. https://repositoriodigital.minam.gob.pe/handle/123456789/178
Ministerio del Ambiente (MINAM). 2016. El Perú y el cambio climático: tercera comunicación nacional del Perú a la Convención Marco de las Naciones Unidas sobre el Cambio Climático. Ministerio del Ambiente, Lima, Peru. https://sinia.minam.gob.pe/documentos/tercera-comunicacion-nacional-peru-convencion-marco-las-naciones
Moomaw, W. R., S. A. Masino, and E. K. Faison. 2019. Intact forests in the United States: proforestation mitigates climate change and serves the greatest good. Frontiers in Forests and Global Change 2:27. https://doi.org/10.3389/ffgc.2019.00027
Myers, N., R. A. Mittermeier, C. G. Mittermeier, G. A. B. da Fonseca, and J. Kent. 2000. Biodiversity hotspots for conservation priorities. Nature 403(6772):853-858. https://doi.org/10.1038/35002501
Ochoa-Tocachi, B. F. 2019. Participatory hydrological monitoring to support sustainable water resources management. Dissertation. Imperial College London, London, UK. https://doi.org/10.25560/75557
Ochoa-Tocachi, B. F., W. Buytaert, B. De Bièvre, R. Célleri, P. Crespo, M. Villacís, C. A. Llerena, L. Acosta, M. Villazón, M. Guallpa, J. Gil-Ríos, P. Fuentes, D. Olaya, P. Viñas, G. Rojas, and S. Arias. 2016. Impacts of land use on the hydrological response of tropical Andean catchments. Hydrological Processes 30(22):4074-4089. https://doi.org/10.1002/hyp.10980
Oliveras, I., Y. Malhi, N. Salinas, V. Huaman, E. Urquiaga-Flores, J. Kala-Mamani, J. A. Quintano-Loaiza, I. Cuba-Torres, N. Lizarraga-Morales, and R.-M. Román-Cuesta. 2014. Changes in forest structure and composition after fire in tropical montane cloud forests near the Andean treeline. Plant Ecology and Diversity 7(1-2):329-340. https://doi.org/10.1080/17550874.2013.816800
Oliveras, I., R. M. Román-Cuesta, E. Urquiaga-Flores, J. A. Quintano Loayza, J. Kala, V. Huamán, N. Lizárraga, G. Sans, K. Quispe, E. Lopez, D. Lopez, I. Cuba Torres, B. J. Enquist, and Y. Malhi. 2018. Fire effects and ecological recovery pathways of tropical montane cloud forests along a time chronosequence. Global Change Biology 24(2):758-772. https://doi.org/10.1111/gcb.13951
Pécastaing, N., and C. Chávez. 2020. The impact of El Niño phenomenon on dry forest-dependent communities’ welfare in the northern coast of Peru. Ecological Economics 178:106820. https://doi.org/10.1016/j.ecolecon.2020.106820
Perry, J., B. Lojka, L. G. Quinones Ruiz, P. Van Damme, J. Houška, and E. Fernandez Cusimamani. 2016. How natural forest conversion affects insect biodiversity in the Peruvian Amazon: Can agroforestry help? Forests 7(4):82. https://doi.org/10.3390/f7040082
Poorter, L., F. Bongers, T. M. Aide, A. M. Almeyda Zambrano, P. Balvanera, J. M. Becknell, V. Boukili, P. H. S. Brancalion, E. N. Broadbent, R. L. Chazdon, D. Craven, J. S. de Almeida-Cortez, G. A. L. Cabral, B. H. J. de Jong, J. S. Denslow, D. H. Dent, S. J. DeWalt, J. M. Dupuy, S. M. Durán, M. M. Espírito-Santo, M. C. Fandino, R. G. César, J. S. Hall, J. L. Hernandez-Stefanoni, C. C. Jakovac, A. B. Junqueira, D. Kennard, S. G. Letcher, J.-C. Licona, M. Lohbeck, E. Marín-Spiotta, M. Martínez-Ramos, P. Massoca, J. A. Meave, R. Mesquita, F. Mora, R. Muñoz, R. Muscarella, Y. R. F. Nunes, S. Ochoa-Gaona, A. A. de Oliveira, E. Orihuela-Belmonte, M. Peña-Claros, E. A. Pérez-García, D. Piotto, J. S. Powers, J. Rodríguez-Velázquez, I. E. Romero-Pérez, J. Ruíz, J. G. Saldarriaga, A. Sanchez-Azofeifa, N. B. Schwartz, M. K. Steininger, N. G. Swenson, M. Toledo, M. Uriarte, M. van Breugel, H. van der Wal, M. D. M. Veloso, H. F. M. Vester, A. Vicentini, I. C. G. Vieira, T. V. Bentos, G. B. Williamson, and D. M. A. Rozendaal. 2016. Biomass resilience of Neotropical secondary forests. Nature 530(7589):211-214. https://doi.org/10.1038/nature16512
Quintero, M., S. Wunder, and R. D. Estrada. 2009. For services rendered? Modeling hydrology and livelihoods in Andean payments for environmental services schemes. Forest Ecology and Management 258(9):1871-1880. https://doi.org/10.1016/j.foreco.2009.04.032
Raboin, M. L., and J. L. Posner. 2012. Pine or pasture? Estimated costs and benefits of land use change in the Peruvian Andes. Mountain Research and Development 32(2):158-168. https://doi.org/10.1659/MRD-JOURNAL-D-10-00099.1
Rasal Sánchez, M., J. Troncos Castro, C. Lizano Durán, O. Parihuamán Granda, D. Quevedo Calle, C. Rojas Idrogo, and G. E. Delgado Paredes. 2011. Características edáficas y composición florística del bosque estacionalmente seco La Menta y Timbes, región Piura, Perú. Ecología Aplicada 10(1-2):61. https://doi.org/10.21704/rea.v10i1-2.414
Román, F., A. Mamani, A. D. Cruz, C. Sandoval, and F. Cuesta. 2018. Orientaciones para la restauración de ecosistemas forestales y otros ecosistemas de vegetación silvestre. Servicio Nacional Forestal y de Fauna Silvestre, Lima, Peru. http://repositorio.serfor.gob.pe/handle/SERFOR/524
Román-Cuesta, R. M., N. Salinas, H. Asbjornsen, I. Oliveras, V. Huaman, Y. Gutiérrez, L. Puelles, J. Kala, D. Yabar, M. Rojas, R. Astete, D. Y. Jordán, M. Silman, R. Mosandl, M. Weber, B. Stimm, S. Günter, T. Knoke, and Y. Malhi. 2011. Implications of fires on carbon budgets in Andean cloud montane forest: the importance of peat soils and tree resprouting. Forest Ecology and Management 261(11):1987-1997. https://doi.org/10.1016/j.foreco.2011.02.025
Ruiz Romero, N. O. 2015. Ensamblaje y dieta de murciélagos (Chiroptera) en la zona noroccidental del Área de Conservación Regional Bosque Seco Salitral - Huarmaca, Piura - Perú. Thesis Universidad Nacional de Piura, Piura, Peru. http://repositorio.unp.edu.pe/handle/UNP/272
Saatchi, S. S., N. L. Harris, S. Brown, M. Lefsky, E. T. A. Mitchard, W. Salas, B. R. Zutta, W. Buermann, S. L. Lewis, S. Hagen, S. Petrova, L. White, M. Silman, and A. Morel. 2011. Benchmark map of forest carbon stocks in tropical regions across three continents. Proceedings of the National Academy of Sciences 108(24):9899-9904. https://doi.org/10.1073/pnas.1019576108
Saatchi, S. S., R. A. Houghton, R. C. Dos Santos Alvalá, J. V. Soares, and Y. Yu. 2007. Distribution of aboveground live biomass in the Amazon basin. Global Change Biology 13(4):816-837. https://doi.org/10.1111/j.1365-2486.2007.01323.x
Scott, D. F., L. A. Bruijnzeel, and J. Mackensen. 2005. The hydrological and soil impacts of forestation in the tropics. Pages 622-651 in M. Bonell and L. A. Bruijnzeel, editors. Forests, water and people in the humid tropics: past, present and future hydrological research for integrated land and water management. Cambridge University Press, Cambridge, UK. https://doi.org/10.1017/CBO9780511535666.032
Seddon, N., A. Chausson, P. Berry, C. A. J. Girardin, A. Smith, and B. Turner. 2020. Understanding the value and limits of nature-based solutions to climate change and other global challenges. Philosophical Transactions of the Royal Society B 375:20190120. https://doi.org/10.1098/rstb.2019.0120
Servicio Nacional Forestal y de Fauna Silvestre (SERFOR). 2018. Experiencias de restauración en el Perú: lecciones aprendidas. SERFOR, Lima, Peru. https://www.serfor.gob.pe/portal/wp-content/uploads/2019/01/Experiencias-de-Restauraci%C3%B3n-en-el-Per%C3%BA-Lecciones-aprendidas.pdf
Servicio Nacional Forestal y de Fauna Silvestre (SERFOR). 2019. Informe del inventario nacional forestal y de fauna silvestre del Perú. SERFOR, Lima, Peru. https://www.serfor.gob.pe/portal/wp-content/uploads/2020/03/INFORME-DEL-INFFS-PANEL-1.pdf
Shukla, P. R., J. Skea, R. Slade, R. van Diemen, E. Haughey, J. Malley, M. Pathak, and J. P. Portugal Pereira, editors. 2019. Technical summary. Pages 35-74 in P. R. Shukla, J. Skea, E. Calvo Buendia, V. Masson-Delmotte, H.-O. Pörtner, D. C. Roberts, P. Zhai, R. Slade, S. Connors, R. van Diemen, M. Ferrat, E. Haughey, S. Luz, S. Neogi, M. Pathak, J. Petzold, J. Portugal Pereira, P. Vyas, E. Huntley, K. Kissick, M. Belkacemi, and J. Malley, editors. Climate change and land: an IPCC special report on climate change, desertification, land degradation, sustainable land management, food security, and greenhouse gas fluxes in terrestrial ecosystems. Intergovernmental Panel on Climate Change, Geneva, Switzerland. https://www.ipcc.ch/site/assets/uploads/sites/4/2020/07/03_Technical-Summary-TS_V2.pdf
Smith, C. C., F. D. B. Espírito-Santo, J. R. Healey, P. J. Young, G. D. Lennox, J. Ferreira, and J. Barlow. 2020. Secondary forests offset less than 10% of deforestation-mediated carbon emissions in the Brazilian Amazon. Global Change Biology 26(12):7006-7020. https://doi.org/10.1111/gcb.15352
Smith, C. C., J. R. Healey, E. Berenguer, P. J. Young, B. Taylor, F. Elias, F. Espírito-Santo, and J. Barlow. 2021. Old-growth forest loss and secondary forest recovery across Amazonian countries. Environmental Research Letters 16(8):085009. https://doi.org/10.1088/1748-9326/ac1701
Socolar, J. B., E. H. Valderrama Sandoval, and D. S. Wilcove. 2019. Overlooked biodiversity loss in tropical smallholder agriculture. Conservation Biology 33(6):1338-1349. https://doi.org/10.1111/cobi.13344
Somers, L. D., J. M. McKenzie, S. C. Zipper, B. G. Mark, P. Lagos, and M. Baraer. 2018. Does hillslope trenching enhance groundwater recharge and baseflow in the Peruvian Andes? Hydrological Processes 32(3):318-331. https://doi.org/10.1002/hyp.11423
Sosa Castillo, J. O. 2016. Valoración económica del secuestro de CO2 en tres tipos de bosque en el distrito del Alto Nanay, Loreto-Perú-2014. Thesis. Universidad Nacional de la Amazonía Peruana, Iquitos, Peru. http://repositorio.unapiquitos.edu.pe/handle/20.500.12737/4283
Soto-Navarro, C., C. Ravilious, A. Arnell, X. de Lamo, M. Harfoot, S. L. L. Hill, O. R. Wearn, M. Santoro, A. Bouvet, S. Mermoz, T. Le Toan, J. Xia, S. Liu, W. Yuan, S. A. Spawn, H. K. Gibbs, S. Ferrier, T. Harwood, R. Alkemade, A. M. Schipper, G. Schmidt-Traub, B. Strassburg, L. Miles, N. D. Burgess, and V. Kapos. 2020. Mapping co-benefits for carbon storage and biodiversity to inform conservation policy and action. Philosophical Transactions of the Royal Society B 375(1794):20190128. https://doi.org/10.1098/rstb.2019.0128
Srinivas, A., and L. P. Koh. 2016. Oil palm expansion drives avifaunal decline in the Pucallpa region of Peruvian Amazonia. Global Ecology and Conservation 7:183-200. https://doi.org/10.1016/j.gecco.2016.06.005
Time, A., M. Garrido, and E. Acevedo. 2018. Water relations and growth response to drought stress of Prosopis tamarugo Phil. A review. Journal of Soil Science and Plant Nutrition 18(2):329-343. https://doi.org/10.4067/S0718-95162018005001103
Timoteo, K., J. Remuzgo, L. Valdivia, F. Sales-Dávila, D. García-Soria, and C. Abanto-Rodriguez. 2016. Estimación del carbono almacenado en tres sistemas agroforestales durante el primer año de instalación en el departamento de Huánuco. Folia Amazónica 25(1):45-54. https://doi.org/10.24841/fa.v25i1.382
Tovar, C., A. C. Seijmonsbergen, and J. F. Duivenvoorden. 2013. Monitoring land use and land cover change in mountain regions: an example in the Jalca grasslands of the Peruvian Andes. Landscape and Urban Planning 112:40-49. https://doi.org/10.1016/j.landurbplan.2012.12.003
Vallet, A., B. Locatelli, H. Levrel, C. B. Pérez, P. Imbach, N. E. Carmona, R. Manlay, and J. Oszwald. 2016. Dynamics of ecosystem services during forest transitions in Reventazón, Costa Rica. Plos One 11(7):e0158615. https://doi.org/10.1371/journal.pone.0158615
van Dijk, A. I. J. M., and R. J. Keenan. 2007. Planted forests and water in perspective. Forest Ecology and Management 251(1-2):1-9. https://doi.org/10.1016/j.foreco.2007.06.010
Vebrova, H., B. Lojka, T. P. Husband, M. E. C. Zans, P. Van Damme, A. Rollo, and M. Kalousova. 2014. Tree diversity in cacao agroforests in San Alejandro, Peruvian Amazon. Agroforestry Systems 88(6):1101-1115. https://doi.org/10.1007/s10457-013-9654-5
Vélez Quesquén, M. A. M., and J. I. Saavedra Chávez. 2019. Estructura del ensamblaje de escarabajos coprófagos (Coleoptera: Scarabaeidae: Scarabaeinae) en un bosque primario y secundario del Bosque de Protección Pagaibamba, Querocoto, Chota, Cajamarca, Perú, 2018. Thesis. Universidad Nacional Pedro Ruiz Gallo, Lambayeque, Peru. https://hdl.handle.net/20.500.12893/5511
Vijay, V., C. D. Reid, M. Finer, C. N. Jenkins, and S. L. Pimm. 2018. Deforestation risks posed by oil palm expansion in the Peruvian Amazon. Environmental Research Letters 13(11):114010. https://doi.org/10.1088/1748-9326/aae540
Villar Cabeza, M. Á., J. E. Cuellar Bautista, and S. L. Valentin Castañeda. 2014. Valoración técnica, económica y ambiental de tres sistemas de silvopasturas en la región Cajamarca. Instituto Nacional de Innovación Agraria, Lima, Peru. http://repositorio.inia.gob.pe/handle/20.500.12955/376
Villogas Ventura, K. E. 2013. Almacenamiento de carbono en sistemas agroforestales (SAF) con cacao (Theobroma cacao L.) en producción. Thesis. Universidad Nacional Agraria de la Selva, Tingo María, Peru. http://repositorio.unas.edu.pe/handle/UNAS/163
Vuille, M., M. Carey, C. Huggel, W. Buytaert, A. Rabatel, D. Jacobsen, A. Soruco, M. Villacis, C. Yarleque, O. E. Timm, T. Condom, N. Salzmann, and J.-E. Sicart. 2018. Rapid decline of snow and ice in the tropical Andes – impacts, uncertainties and challenges ahead. Earth-Science Reviews 176:195-213. https://doi.org/10.1016/j.earscirev.2017.09.019
Vuille, M., B. Francou, P. Wagnon, I. Juen, G. Kaser, B. G. Mark, and R. S. Bradley. 2008. Climate change and tropical Andean glaciers: past, present and future. Earth-Science Reviews 89(3-4):79-96. https://doi.org/10.1016/j.earscirev.2008.04.002
Walentowski, H., S. Heinrichs, S. Hohnwald, A. Wiegand, H. Heinen, M. Thren, O. A. Gamarra Torres, A. B. Sabogal, and S. Zerbe. 2018. Vegetation succession on degraded sites in the Pomacochas basin (Amazonas, N Peru)—ecological options for forest restoration. Sustainability 10(3):609. https://doi.org/10.3390/su10030609
Whaley, O. Q., D. G. Beresford-Jones, W. Milliken, A. Orellana, A. Smyk, and J. Leguía. 2010. An ecosystem approach to restoration and sustainable management of dry forest in southern Peru. Kew Bulletin 65(4):613-641. https://doi.org/10.1007/s12225-010-9235-y
Whitworth, A., R. Downie, R. von May, J. Villacampa, and R. MacLeod. 2016. How much potential biodiversity and conservation value can a regenerating rainforest provide? A ‘best-case scenario’ approach from the Peruvian Amazon. Tropical Conservation Science 9(1):224-245. https://doi.org/10.1177/194008291600900112
Wilson, S. J., J. Schelhas, R. Grau, A. S. Nanni, and S. Sloan. 2017. Forest ecosystem-service transitions: the ecological dimensions of the forest transition. Ecology and Society 22(4):38. https://doi.org/10.5751/ES-09615-220438
Young, K. R., and B. León. 2007. Tree-line changes along the Andes: implications of spatial patterns and dynamics. Philosophical Transactions of the Royal Society B 362(1478):263-272. https://doi.org/10.1098/rstb.2006.1986
Zamorano-Elgueta, C., J. M. R. Benayas, L. Cayuela, S. Hantson, and D. Armenteras. 2015. Native forest replacement by exotic plantations in southern Chile (1985–2011) and partial compensation by natural regeneration. Forest Ecology and Management 345:10-20. https://doi.org/10.1016/j.foreco.2015.02.025
Zavala, W., E. Merino, and P. Peláez. 2018. Influencia de tres sistemas agroforestales del cultivo de cacao en la captura y almacenamiento de carbono. Scientia Agropecuaria 9(4):493-501. https://doi.org/10.17268/sci.agropecu.2018.04.04
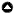
Fig. 1

Fig. 1. (A) Map of Peru showing the coastal, Andean, and Amazonian regions. Photographs showing mature forest (B), secondary forest regrowth following tea plantation (C), and a tea agroforestry system (D), all located in the Huayopata District, Cuzco region (indicated by an asterisk in panel A). (E) Photograph of coastal dry secondary forest located in the Mangamanguilla Private Conservation Area, Piura region, in northwestern Peru (indicated by a triangle in panel A).
Fig. 2

Fig. 2. (a) Carbon stocks and (b) biodiversity values in secondary forests and in two human-assisted post-deforestation regrowth forests (tree plantations and agroforestry system) in the Amazon region. In Fig. 2b, percent of species represents a percentage compared to a mature forest baseline. MF: mature forest. Data used for these figures are in the Table A2.
Fig. 3

Fig. 3. Carbon stocks and (b) biodiversity values in secondary forests and in two human-assisted post-deforestation regrowth forests (tree plantations and agroforestry system) in the Andean region. In Fig. 3b, percent of species represents a percentage compared to a mature forest baseline. MF: mature forest. Data used for these figures are in the Table A2.
Fig. 4

Fig. 4. (A) Framework showing the relationship between the forest transitions, their relative ecological outcomes, and their drivers, linking the environmental and socioeconomic systems. Forest transition is defined as a long period of decline in forest area, superseded by forest recovery. The “+” and “−” represent the potential amount of each service that would be expected to return depending on the forest transition type. (B) The relative importance of key ecosystem services provided by secondary forests in Amazonian, Andean, and coastal regions.