|
|
|
Copyright © 2003 by the author(s). Published here under license by The Resilience Alliance.
Go to the pdf version of this article.
The following is the established format for referencing this article:
Read, A. J. and C. R. Brownstein. 2003. Considering other consumers:fisheries, predators, and Atlantic herring in the Gulf of Maine. Conservation Ecology 7(1): 2. [online] URL: http://www.consecol.org/vol7/iss1/art2/
Report Considering Other Consumers: Fisheries, Predators, and Atlantic Herring in the Gulf of Maine Andrew J. Read and Carrie R. Brownstein
Duke University
- Abstract
- Introduction
- Methods
- Results
- Discussion
- Conclusions
- Responses to This Article
- Acknowledgments
- Literature Cited
After decades of overexploitation and severe depletion, Atlantic herring stocks in waters of the northeastern United States have recovered. Fishery managers now consider the herring resource to be underexploited. Nevertheless, some fishery managers and sustainable fishery advocates in New England have expressed concern that the fishery management plan may not adequately consider the importance of herring as prey for marine mammals, seabirds, and piscivorous fish. Several studies suggest that consumption by these predators is significant, yet trophic interactions are not explicitly considered in stock assessment models. Instead, as in most fisheries stock assessments, predation is subsumed within the natural mortality rate, and no empirical estimates of herring consumption are used in the models. The goal of the present study was to assess the consumption of herring by marine mammals and to compare this level of consumption with estimates of natural mortality derived from herring stock assessment models. Using the most recent estimates of abundance and the best available data on diet, we estimated total annual consumption of herring by eight marine mammal species in the Gulf of Maine. Our results indicate that marine mammals consume 93,802–189,898 metric tons (mt; 1 metric ton = 1000 kg) of herring annually. In addition, piscivorous fish and seabirds are important predators of herring. We estimate that the consumption of herring by these upper trophic level predators may have exceeded the estimate of natural mortality used in stock assessment models by more than fourfold in 1991. We suggest that fisheries management must move beyond a single-species approach to one that includes formal consideration of trophic relationships.
KEY WORDS: Atlantic herring, ecosystem management, fisheries management, Gulf of Maine, marine mammals, piscivores, protected species, single-species approach, stock assessment, trophic interactions.
Published: January 20, 2003
Managing the extraction of natural resources to account for larger ecosystem dynamics is a challenging task for both terrestrial and aquatic natural resource managers. Whether concerned with fish or forests, managers seek to obtain optimum benefits from resource extraction while considering biological and ecological processes such as reproduction, regeneration, predation, and competition. In some systems, however, the data required to adequately understand ecosystem processes may be difficult to obtain or unavailable. Yet, despite this difficulty, federal legislation may mandate that resources are managed to protect additional species and habitats.
Efforts to model ecosystem bioenergetics are underway in many resource sectors. Approaches such as Ecopath offer a method for scientists to understand ecosystem processes such as feeding interactions or nutrient flow in the context of various harvest regimes.
Fishery managers face the dual responsibility of obtaining the optimum yield from fisheries while simultaneously protecting certain other species (Gerber et al. 1999). The bycatch of protected species such as sea turtles and marine mammals is well recognized (Crowder and Murawski 1998, National Research Council 1999), but the ecological role of commercially important fish species as prey of protected species has received less attention in the management process (Overholtz et al. 2000). Most commercial fisheries in the United States are managed on a single-species basis, but trophic interactions occur within a complex, multispecies environment.
In the Gulf of Maine, Atlantic herring (Clupea harengus) is both a commercially valuable species and an important prey species for a large number of marine mammals, seabirds, and piscivorous fish (Caddy and Iles 1973, Overholtz et al. 1991, 2000, NEFMC 1999). The herring resource in the Gulf of Maine is currently considered to be underexploited by the New England Fishery Management Council. In 2001, the ex-vessel value of the herring fishery was U.S. $15.6 million, with landings of 118,000 mt (NEFMC 2002). Landings and the value of the fishery have increased since 1986, when the fishery was valued at only U.S. $4.3 million. Markets currently include canned and frozen herring for human consumption and bait for the lobster fishery. Because of the current abundance of herring, there is commercial interest in further expansion of the fishery (NEFMC 2002).
Although managers consider the herring fishery to be underexploited today, this has not always been the case. Growth of the herring fishery began in the 1870s in eastern Maine, with the development of the sardine canning industry and the lobster fishery. Other markets included use of herring for fertilizer, for smoking and pickling, for fresh herring, and for reduction purposes. Landings of herring reached 80,000–90,000 mt by the end of the 19th century and fluctuated throughout the first half of the 20th century. In the 1960s, however, overfishing by distant-water fleets resulted in a collapse of the Georges Bank stock (Friedland 1998). With the passage of the Magnuson Fishery Conservation and Management Act of 1976–1977, distant-water fleets were largely excluded from U.S. waters, and abundance of the overall stock complex eventually increased under relatively light fishing pressure. In 1997, the year for which the most recent stock assessment was conducted, the total herring biomass was estimated at 2.9 million mt, with a spawning stock biomass of approximately 1.8 million mt (Fig. 1). Although several spawning stocks contribute to the herring complex, herring stocks in the Gulf of Maine and Georges Bank today are assessed as a single coastal complex (NEFMC 2002).
Management plans for the Atlantic herring fishery (see Fig. 2 for an illustration of management areas; Areas 1A and 1B represent the Gulf of Maine) are developed jointly by the New England Fisheries Management Council (NEFMC) and the Atlantic States Marine Fisheries Commission (ASMFC), and are approved by the National Marine Fisheries Service (NMFS) and the ASMFC. Within the herring Fishery Management Plan (FMP), trophic interactions between herring and predators such as marine mammals, seabirds, and piscivorous fish are subsumed within the natural mortality estimates used in stock assessment models. Because no empirical data for predation are incorporated in the models, trophic relationships are considered implicitly in the assumed natural mortality rate of 0.2 rather than explicitly (T. Nies, NEFMC, personal communication). Fishery scientists use these stock assessment models to estimate the Maximum Sustainable Yield for the herring fishery, which is subsequently applied by managers to determine Optimum Yield and Total Allowable Catch.
Fishery managers and sustainable fishery advocates in New England have expressed concern about the lack of quantitative data regarding the role of herring in relation to other consumers within the ecosystem (M. Pentony, NEFMC, personal communication; N. Dorry, Greenpeace, personal communication). Currently, there is a lack of guidance within fisheries policy to direct managers to explicitly address such trophic relationships in FMPs. Despite this lack of guidance, fishery managers are mandated by the Marine Mammal Protection Act (MMPA) and the Endangered Species Act (ESA) to protect certain species that interact ecologically with commercial fish species.
As required by Section 7 of the Endangered Species Act, in September 1999 the Office of Protected Species within NMFS issued a biological opinion assessing potential effects of the herring fishery on threatened or endangered species. The report concluded that “the proposed herring fishery is not likely to jeopardize the continued existence of threatened or endangered species or designated critical habitat.” This consultation reports, however, that new information on prey requirements for endangered whales may require re-initiation of the consultation (NMFS 1999a).
Understanding the prey requirements of upper trophic level predators requires information on their diet, abundance, energetic requirements, and functional responses to variation in prey density. Despite the complex nature of these trophic interactions, some researchers have attempted to investigate the relationships between marine mammals and their prey in New England. For example, Kenney et al. (1997) estimated consumption of finfish, squid, and zooplankton by cetaceans in the Northeast Shelf ecosystem and concluded that cetaceans are significant consumers of prey. Overholtz et al. (1991) examined predation on herring by four species of predatory fish, 10 species of marine mammals, and three species of seabirds off the northeastern United States. Overholtz et al. (1991) estimated that these predators consumed approximately 30,000 mt of herring per year, with marine mammals accounting for 19,300 mt per year from 1988 to 1992. The herring FMP refers to these estimates and concludes that 30,000 mt may be an underestimate, but further notes that even annual consumption of 50,000 mt would represent only 2.5% of the 1990 total herring stock size (NEFMC 1999). More recently, Overholtz et al. (2000) estimated the consumption of herring and other pelagic fish and squid by 12 species of predatory fish in New England and concluded that these piscivorous fishes could impact the demography of these prey stocks in a significant fashion.
Kenney et al. (1997) and Overholtz et al. (1991) demonstrated that marine mammals are significant consumers of herring and other species in New England, but in the present analysis, we explore the possibility that current levels of herring consumption by marine mammals may be even higher. Both Kenney et al. (1997) and Overholtz et al. (1991) generated consumption estimates using marine mammal abundance data from the University of Rhode Island’s aerial surveys conducted from 1979 to 1982 and 1980–1983 shipboard surveys by the Manomet Bird Observatory. Since those papers were published, however, the National Marine Fisheries Service has generated new and more precise estimates of marine mammal abundance. The recent abundance estimates have also incorporated changes in population size for some species. For example, the population of harbor seals (Phoca vitulina) on the Maine coast increased from a minimum of 10,540 in 1981 (Gilbert and Stein 1981) to 30,990 in 1997 (Gilbert and Guldager 1998). Also, the abundance of minke whales (Balaenoptera acutorostra) ranging from Cape Hatteras to Nova Scotia during the years 1978–1982 was estimated to be 320 individuals (CETAP 1982). In 1991–1992, the population estimate for the Gulf of Maine and the Bay of Fundy was 2650 individuals (Palka 1995). These population changes may reflect either population growth or improvements in survey techniques, or both.
We use abundance estimates for marine mammals in the Gulf of Maine, generated between 1991 and 1997 (NMFS 1999b), to calculate estimates of the consumption of herring. We then compare these figures with estimates of the natural mortality of herring derived from stock assessment models. Our study is an effort to better understand predation by eight marine mammal species for which herring constitutes a significant portion of their diet in the Gulf of Maine: fin whale (Balaenoptera physalus), minke whale, humpback whale (Megaptera novaeangliae), pilot whale (Globicephala melas), harbor porpoise (Phocoena phocoena), white-sided dolphin (Lagenorhynchus acutus), harbor seal, and gray seal (Halichoerus grypus). The ultimate goal of our research is to support a transition to a fisheries management system in which trophic relationships are directly incorporated into the setting of fishing quotas.
Using information available in the literature, we estimated herring consumption by the eight marine mammal predators just noted. First, we used an allometric equation from Innes et al. (1987) to calculate daily food consumption by each marine mammal species. Innes et al. used this equation to conduct interspecific comparisons over a wide range of species and found that, under standardized conditions, rates of food consumption by an individual vary in relation to its body mass:
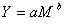 |
(1) |
where Y is consumption (in kilograms per day) and M is body mass (in kilograms). Sources of data on body mass are presented in Appendix 1. For marine mammals, a is estimated to be 0.123 and b is estimated to be 0.80 (Innes et al. 1987). Thus, this equation can be written as:
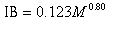 |
(2) |
where IB is ingested biomass (in kilograms per day).
This equation can be transformed to a linear form:
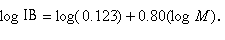 |
(3) |
Second, using the coefficients of variation for abundance estimates from marine mammal stock assessment reports (NMFS 1999b), we generated high, best, and low estimates of population size by adding and subtracting one standard deviation to and from the best estimate of abundance. These estimates of abundance were generated from surveys in the Gulf of Maine conducted between 1991 and 1997 (see Appendix 1). These estimates were then multiplied by the individual daily consumption previously calculated to determine high, best, and low estimates of daily consumption (in kilograms per day) by the entire population in the Gulf of Maine. Third, we calculated annual consumption in metric tons (mt; 1 mt = 1000 kg) for each population. To do this, we multiplied population consumption by the number of days that each species spent in the Gulf of Maine. We assumed that time spent for a particular species was either 182 or 365 days, depending on whether a species was a resident of the region or migratory. Fourth, we multiplied this value by the proportion of the species’ diet (see Appendix 1 for data sources) that consisted of herring to obtain the total annual herring consumption (in metric tons) for each species. Our calculations and results appear in Table 1. Consistent with Kenney et al. (1997), we accounted for statistical variability only within the abundance estimates, because variances were not consistently available from other data.
The estimated total annual consumption of Atlantic herring by all eight marine mammal species ranged from 93,802 to 189,898 metric tons (mt), using the low and high estimates of abundance respectively (Table 1). Using the best estimates of population size, we estimated total annual consumption as 141,341 mt. It is important to note that most estimates of the abundance of marine mammals were generated prior to 1997 and that many of these populations are growing; hence, this estimate of predation is likely to be negatively biased.
Using Virtual Population Analysis (VPA) models, fishery scientists predict the total biomass of herring from historical estimates of the size and average biomass of all age classes, derived from fish catches and assumptions regarding the rates of natural mortality. As is the case with many other commercially valuable pelagic species, this approach assumes an instantaneous natural mortality rate (m) of 0.2 for all age classes of Atlantic herring (NEFSC 1998a). For the purpose of comparison with consumption estimates from this study, we converted the total amount (in numbers) of herring dying (as presented in the stock assessment) to the total mass of herring dying from natural mortality. To do this, we first applied the herring abundance estimates from the VPA and an assumed natural mortality rate of 0.2 to the equation:
 |
(4) |
to calculate the total biomass in numbers of herring dying of natural mortality in each age class from 1991 to 1997. In this analysis, N0 refers to stock abundance in the current year, m is the instantaneous rate of natural mortality, f is the instantaneous rate of fishing mortality, and z is the instantaneous rate of total mortality. We then used the average mass per individual in each age class (also taken from the fishery landings) to determine the mass of fish that died every year in each age class. Finally, we summed the mass of fish that died in all age classes to get a total mass of herring that died from natural mortality each year from 1991 to 1997. The results of this analysis indicate that from 90,183 mt to 672,555 mt (mean 321,996 mt) of herring died annually from natural mortality during this period (Table 2). This value encompasses natural mortality from all sources of predation including piscivorous fish, marine mammals, and seabirds. In comparison, our estimates predict that eight marine mammal species consume from 93,802 mt to 189,898 mt of herring annually.
Our lowest estimate of total consumption of herring by marine mammals from 1991 to 1997 (93,802 mt) is larger than the instantaneous rate of natural mortality assumed by the VPA in 1991 (90,183 mt). Because our estimate pertains only to consumption by marine mammals in the Gulf of Maine—and not throughout the entire range of the herring stock—it is likely a conservative estimate. When we also consider the estimates of consumption by piscivorous fish (273,000 mt in 1991) generated by Overholtz et al. (2000), we find that predation on herring by fish and mammals exceeded the level of natural mortality assumed in the VPA by fourfold. Estimates of predation by piscivorous fish alone were over three times the natural mortality assumed in the VPA. Thus, our findings highlight a discrepancy between direct estimates of consumption and the implicit consideration of consumption from the assumption of a constant natural mortality rate across all age classes in the VPA. The VPA assumes a constant value of M across all years and age classes, but in reality, the intensity of mortality due to predation will vary with predator abundance and other factors. For example, we know that some marine mammal populations are increasing in abundance in the Gulf of Maine. Humpback whale and harbor seal populations are known to be increasing and gray seal populations are considered likely to be increasing (NMFS 1999b). Because all marine mammals in U.S. waters are protected under the Marine Mammal Protection Act of 1972—and some species such as fin and humpback whales are endangered and receive added protection under the Endangered Species Act of 1973—, the abundance of many marine mammal populations in the Gulf of Maine is increasing or is likely to increase in the future (Read and Wade 2000). Because herring are an important prey item, consumption by these predators will probably increase as marine mammal populations grow.
During the time period covered by this study (1991–1997), many populations of important fish predators declined. Overholtz et al. (2000) showed that consumption of herring by piscivores declined in the late 1990s, following declines in abundance of Georges Bank cod (Gadus morhua), spiny dogfish (Squalus acanthias), silver hake and white hake (Urophycis spp.), and bluefish (Pomatomus saltatrix). Unfortunately, we do not have comparable annual estimates of marine mammal abundance and consumption, although, as previously noted, we believe that most of these mammalian predators were increasing during the study period.
Thus, in some years (1991 and 1992), natural mortality of herring may have been underestimated to a significant extent, even considering only the consumption by marine mammals and piscivorous fish. Fortunately, this did not have adverse effects on herring stocks, which have continued to grow under light fishing pressure and the removal of much of the biomass of fish predators. Nevertheless, we believe that the assumption of a constant natural mortality rate, despite changes in predator abundance, could lead managers to authorize a Total Allowable Catch (TAC) for the fishery that is insufficient to sustain the fishery, while also assuring a continued supply of prey to upper trophic level predators.
For example, the Sustainable Fisheries Act of 1996 requires that fishery managers in New England implement recovery plans to rebuild overfished populations, so it is likely that groundfish populations will increase in the future and that their predation on herring will also increase. At the same time, populations of marine mammals, protected under the MMPA and ESA, will continue to grow in the Gulf of Maine. In addition, it is likely that the fishery for herring will expand, particularly given current restrictions on other traditional fisheries. Thus, although herring stocks are currently robust, they will face increased pressure from both predators and humans in the near future. It is unclear whether a traditional, single-species management approach will be sufficient to balance the competing needs of recovering groundfish, expanding populations of marine mammals, and the herring fishery. In particular, the simplistic assumption of a constant natural mortality rate of 0.2 is unlikely to capture the complex trophic dynamics of this system.
Under current laws governing U.S. fisheries management, managers are required to consider the health of the marine environment (not solely the fishery resource) when designing their plans. Simultaneously, under the MMPA and the ESA, managers are legally obligated to protect marine mammals and endangered species. Nowhere within the Magnuson-Stevens Fishery Conservation and Management Act (MSFCMA), the law that governs fisheries in federal waters of the USA, are managers told how this should be done. One solution for formal consideration of trophic relationships under the current management approach is to incorporate quantitative estimates of predation into the Optimum Yield (OY) setting process. Estimates of consumption could be considered “ecological factors” and could serve as points for adjustment in estimating OY and TAC. Already, the definition of OY refers specifically to the protection of marine ecosystems and the consideration of ecological factors. The first two points in the definition of Optimum Yield are: “the amount of fishing which (1) will provide the greatest overall benefit to the Nation, particularly with respect to food production and recreational opportunities, and taking into account the protection of marine ecosystems, and (2) is prescribed as such on the basis of maximum sustained yield from a fishery, as reduced by any relevant economic, social, or ecological factor” (MSFCMA, Sec. 3(28), 16 U.S.C. 1802).
Ecological interactions: the significance of reduced prey availabilityA consideration of the relationship between predation and resource extraction for human use is applicable to all areas of wildlife conservation. In the present study, by estimating the consumption of herring by marine mammals, we aimed to quantitatively assess the importance of herring as a prey species for marine mammal predators in the Gulf of Maine. Understanding the significance of consumption by these large predators and by other predators such as piscivorous fish is critical for determining whether current levels of resource extraction are appropriate. Likewise, it is also critical for understanding the risk of reduced fishing due to reduced herring availability, should the abundance of all predator groups increase substantially. Unfortunately, predicting the impact of changes in predator abundance and in prey availability requires knowledge of complex biological interactions between herring and marine ecosystems. For example, it is interesting to note that stocks of herring have increased over the past decade as populations of predatory fishes have been depleted. However, at the same time, populations of marine mammals have increased. With our current limited understanding of the trophic dynamics of this system, it is not possible to fully evaluate the relative roles of mammalian and fish predators in affecting the abundance of herring stocks. It would be presumptuous to assume that we understand these interactions fully, but some insight can be gained from related studies that demonstrate the potential effects of changing prey availability upon marine mammal populations and of changing levels of consumption with fluctuations in prey populations.
Weinrich et al. (1997) and Payne et al. (1990) described shifts in the distribution of humpback whales in the Gulf of Maine in response to the collapse of herring stocks in the 1970s. As herring declined, populations of sand lance (Ammodytes spp.; a competitor of herring) exploded, and humpbacks in the Gulf of Maine moved to areas with a greater abundance of sand lance. Weinrich et al. (1997) also documented sharp declines and the eventual abandonment of Stellwagen Bank by humpbacks in 1994, and concluded that changes in the abundance of herring may have led to major shifts in the distribution of humpback whales in the Gulf of Maine. Although very little literature exists describing the implications of prey shifting on marine mammals, Lavigne (1995) suggested that prey shifting may be less common than many scientists have assumed. “To date,” Lavigne (1995) stated, “we usually assume that marine mammals, particularly seals, are opportunistic predators, consuming those prey that happen to be most abundant in a particular place at a particular time. This perception leads to the belief that when a particular prey becomes less abundant, it will have little effect because the marine mammals predator will simply switch to another, more available prey. This is not always the case.” For example, the condition of harp seal populations declined when capelin (Mallotus villosus) abundance decreased in the Northwest Atlantic during the 1970’s. This effect suggests that the seals were not able to switch prey (Stewart and Lavigne 1984).
In addition, some scientists have considered the role that marine mammals can play in the recovery of a fish population. Sissenwine et al. (1984) explained that the ability of a fish population to recover from a decline is not only dependent on reduced catch levels from a fishery, but also may depend on predation due to marine mammals. “Like a fishery,” Sissenwine et al. (1984) state, “marine mammals have a potential for density dependent population effects. If a fishery, or marine mammals, switch to a more abundant target as a fish population becomes rare, then the system has the potential for compensation. On the other hand, if the fishery, or marine mammals, are able to remove a nearly constant amount of fish, even as fish abundance decreases, the system is potentially depensatory.” Sissenwine et al. (1984) suggested that depensation (or decrease in production per unit biomass) may explain why the Georges Bank herring population failed to recover despite reductions in catch in 1976.
Potential sources of errorOur estimates of consumption of herring by marine mammals include several potential sources of error. For example, the marine mammal abundance surveys include coefficients of variation ranging from 0.067 for the humpback whale to higher values for species such as white-sided dolphins (0.63) and fin whales (0.59) (NMFS 1999b). The allometric equation is also associated with some degree of uncertainty. This equation estimates consumption from body size, but does not account for variation in body size within a species because the values of body mass used were averaged for each species. Even though the estimates of consumption from this equation are likely to be imprecise, there is no reason to believe that they are biased. Also, by using diet data obtained from different sources, variation is introduced into estimates of the proportion of each species’ diet that consists of herring. For example, data on the diet of pilot whales and harbor porpoises were obtained from large samples of specimens taken as bycatch and are likely to be more accurate than estimates of the diet of humpback and fin whales, which were taken from the Overholtz et al. (1991) study. Simplifying the time that each species spent in the Gulf of Maine to 182 or 365 days may introduce additional imprecision. Katona et al. (1993) estimated that migratory fin and minke whales spend seven and eight months (respectively) in the Gulf of Maine. If the estimates of Katona et al. are correct, then perhaps our estimates are overly conservative.
To compare our estimates of consumption to the natural mortality rate, we used the VPA’s fishery-dependent data to examine the proportion of herring caught from each age class. In doing so, we assumed that the age composition of the fishery was the same as that of the herring population. Fishery-dependent sampling techniques potentially can incorporate bias, because fishermen may target specific age classes for harvest, under-report their catches, or high-grade. (High-grading was defined by Gillis et al. (1995) as the discarding of catch, before a vessel’s capacity is filled,to meet regulations or physical catch limits). The New England Fishery Management Council, however, has not had any reports of high-grading (T. Nies, NEFMC, personal commnunication).
Finally, we assumed that marine mammals do not select certain age classes of herring as prey; thus predation reflected the age structure of the population. This is probably not the case and predation by marine mammals may be more intense on younger juvenile herring than on older fish (e.g., Gannon et al. 1998). Estimates of predation on walleye pollock (Theragra chalcogramma) in the eastern Bering Sea using multispecies VPAs suggest that predation may be particuarly intense in young year classes, the biomass of which may not be well captured by traditional VPA models (Livingston and Jurad-Molina 2000).
We recommend an approach that includes a mechanism for fisheries management to move more progressively toward formal consideration of trophic relationships. We also suggest a research agenda incorporating estimates of consumption of Atlantic herring by major predators into ecosystem modeling approaches such as Ecopath and Ecosim. The intention of these suggestions is to encourage fishery managers to move beyond a single-species approach to management of our public fishery resources. Due to the current abundance of the herring population, managers have a unique opportunity to manage proactively. Although achieving such objectives is by no means simple, such a transition is critical if we are to maintain healthy marine ecosystems.
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a comment, follow this link. To read comments already accepted, follow this link.
This research was supported by several individuals at the Duke University Marine Lab in Beaufort, North Carolina, USA. In particular, we thank Kevin Craig for his expertise in fisheries science and conversations regarding natural mortality rates, as well as with editing. Dr. Larry Crowder provided thoughtful input into the ecological component of the project and Dr. Michael Orbach offered insightful comment into the policy component. Tom Nies of the New England Fishery Management Council answered many questions about the status of the herring fishery. Finally, we thank the reviewers for their thoughtful reviews and helpful suggestions, which have greatly improved this manuscript.
Boulva, J., and I. A. McLaren. 1979. Biology of the harbor seal in eastern Canada. Bulletin 200 of the Fisheries Research Board of Canada. Government of Canada Fisheries and Oceans, Ottawa, Canada.
Bowen, W. D., J. W. Lawson, and B. Beck. 1993. Seasonal and geographic variation in the species composition and size of prey consumed by grey seals (Halichoerus grypus) on the Scotian Shelf. Canadian Journal of Fisheries and Aquatic Sciences 50:1768–1778.
Caddy, J. F., and T. D. Iles. 1973. Underwater observations on herring spawning grounds on Georges Bank. International Commission on North American Fisheries Research Bulletin 10:131–139.
CETAP. 1982. A characterization of marine mammals and turtles in the mid-and north Atlantic areas of the U.S. outer continental shelf. Cetacean and Turtle Assessment Program, University of Rhode Island. Final Report #AA551-CT8-48 to the Bureau of Land Management, Washington, D.C., USA.
Crowder, L., and S. Murawski. 1998. Fisheries bycatch: implications for management. Fisheries 23:8–17.
Ferland, A. 1999. Feeding habits of harbor seals along the coast of Cape Cod, Massachusetts and the Northern Gulf of Maine. Thesis. Boston University, Boston, Massachusetts, USA.
Friedland, K. 1998. Status of fisheries resources off the Northeastern United States. NOAA [National Oceanic and Atmospheric Administration] Technical Memorandum NMFS-NE-115. Resource Evaluation and Assessment Division, Northeast Fisheries Science Center. [Online, URL: http://www.nefsc.noaa.gov/sos/spsyn/pp/herring/.]
Gannon, D. P., J. E. Craddock, and A. J. Read. 1998. Autumn food habits of harbor porpoises, Phocoena phocoena, in the Gulf of Maine. Fishery Bulletin 96:428–437.
Gannon, D. P., A. J. Read, J. E. Craddock, K. M. Fristrup, and J. R. Nicholas. 1997. Feeding ecology of long-finned pilot whales Globicephala melas in the western North Atlantic. Marine Ecology Progress Series 148:1–10.
Gerber, L. R., W. S. Wooster, D. P. DeMaster, and G. R. Van Blaricom. 1999. Marine mammals: new objectives in U.S. fishery management. Reviews in Fisheries Science 71:23–38.
Gilbert, J. R., and N. Guldager. 1998. Status of harbor and gray seal populations in northern New England. Final Report to the National Marine Fisheries Service, Northeast Fisheries Science Center, Woods Hole, Massachusetts, USA. Under NMFS/NER Cooperative Agreement 14-16-0091557.
Gilbert, J. R., and J. L. Stein. 1981. Harbor seal populations and marine mammal fisheries interactions. 1981 Annual Report, Contract NA-80-FA-C-00029. Northeast Fisheries Science Center, National Marine Fisheries Service, Woods Hole, Massachusetts, USA.
Gillis, D. M., E. K. Pikitch, and R. M. Peterman. 1995. Dynamic discarding decisions: foraging theory for high-grading in a trawl fishery. Behavioral Ecology 6:146–154.
Innes, S., D. M. Lavigne, W. M. Earle, and K. M. Kovacs. 1987. Feeding rates of seals and whales. Journal of Animal Ecology 56:115–130.
Katona, S. K., V. Rough, and D. T. Richardson. 1993. Guide to whales, porpoises and seals from Cape Cod to Newfoundland. Fourth edition, revised. Smithsonian Institution Press, Washington, D.C., USA.
Kenney, R. D., M. A. M. Hyman, and H. E. Winn. 1985. Calculation of standing stocks and energetic requirements of the cetaceans of the Northeast United States Outer Continental Shelf. NOAA [National Oceanic and Atmospheric Administration] Technical Memorandum NMFS-F/NEC-41. U.S. Department of Commerce, Woods Hole, Massachusetts, USA.
Kenney, R. D., G. P. Scott, T. J. Thompson, and H. E. Winn. 1997. Estimates of prey consumption and trophic impacts of cetaceans in the USA northeast continental shelf ecosystem. Journal of Northwest Atlantic Fishery Science 22:155–171.
Lavigne, D. M. 1995. Ecological interactions between marine mammals, commercial fisheries and their prey: unraveling the tangled web. Technical Report Number 95-02, International Marine Mammal Association, Guelph, Ontario, Canada.
Livingston, P. A., and J. Jurado-Molina. 2000. A multispecies virtual population analysis of the eastern Bering Sea. ICES Journal of Marine Science 57:294–299.
Mohn, R., and W. D. Bowen. 1996. Grey seal predation on the Eastern Scotian Shelf: modeling the impact on Atlantic cod. Canadian Journal of Fisheries and Aquatic Sciences 53:2722–2738.
NMFS (National Marine Fisheries Service). 1999a. Office of Protected Species ESA [Endangered Species Act] Section 7 Consultation on Federal Atlantic Herring Fishery Management Plan. National Marine Fisheries Service, Silver Spring, Maryland, USA.
NMFS (National Marine Fisheries Service). 1999b. Northeast Region. U.S. Atlantic Marine Mammal Stock Assessments—1998. NOAA Technical Memorandum NMFS-3NE-116. National Marine Fisheries Service, Woods Hole, Massachusetts, USA.
National Research Council. 1999. Sustaining marine fisheries. Committee on Ecosystem Management for Sustainable Marine Fisheries. National Academy, Washington, D.C., USA.
NEFMC (New England Fisheries Management Council). 1999. Atlantic Herring Fishery Management Plan and Environmental Impact Statement. New England Fisheries Management Council, Newburyport, Massachusetts, USA.
NEFMC (New England Fisheries Management Council). 2002. Atlantic Herring Stock Assessment and Fishery Evaluation Report for the 2001 Fishing Year. New England Fisheries Management Council, Newburyport, Massachusetts, USA.
Overholtz, W. J., J. S. Link, and J. E. Suslowicz. 2000. Consumption of important pelagic fish and squid by predatory fish in the northeastern USA shelf ecosystem with some fishery comparisons. ICES Journal of Marine Science 57:1147–1159.
Overholtz, W. J., S. A. Murawski, and K. L. Foster. 1991. Impact of predatory fish, marine mammals, and seabirds on the pelagic fish ecosystem of the northeastern USA. International Council for Exploration of the Sea Marine Science Symposium 193:198–208.
Palka, D. 1995. Abundance estimate of the Gulf of Maine harbor porpoise. Pages 27–50 in A. Bjørge and G. P. Donovan, editors. Biology of Phocoenids. Reports of the International Whaling Commission Special Issue 16.
Palka, D., A. J. Read, and C. W. Potter. 1997. Summary of knowledge of white-sided dolphins (Lagenorhynchus acutus) from U.S. and Canadian Atlantic waters. Reports of the International Whaling Commission 47:729–734.
Payne, P. M., D. N. Wiley, S. B. Young, S. Pittman, P. J. Clapham, and J. W. Jossi. 1990. Recent fluctuations in the abundance of baleen whales in the southern Gulf of Maine in relation to changes in prey. Fishery Bulletin (U.S.) 88:687–696.
Read, A. J., and P. R. Wade. 2000. Status of marine mammals in the United States. Conservation Biology 14:929–940.
Recchia, C. A., and A. J. Read. 1988. Stomach contents of harbour porpoises, Phocoena phocoena (L.), from the Bay of Fundy. Canadian Journal of Zoology 67:2140–2146.
Sissenwine, M. P., W. J. Overholtz, and S. H. Clark. 1984. In search of density dependence. Proceedings of the Workshop on Biological Interactions among Marine Mammals and Commercial Fisheries in the southeastern Bering Sea. Alaska Sea Grant Report 84-1.
Stewart, R. E. A., and D. M. Lavigne. 1984. Energy transfer and female condition in nursing harp seals Phoca groenlandica. Holarctic Ecology 7:182–194.
Weinrich, M., M. Martin, R. Griffiths, J. Bove, and M. Schilling. 1997. A shift in distribution of humpback whales, Megaptera novaeangliae, in response to prey in the southern Gulf of Maine. Fishery Bulletin 95:826–836.
Address of Correspondent:
Andrew J. Read
Duke University Marine Lab
135 Duke Marine Lab Road
Beaufort, NC 28516-9721 USA
Phone: 252-504-7590
aread@duke.edu
![]()