|
|
|
Copyright © 2003 by the author(s). Published here under license by The Resilience Alliance.
Go to the pdf version of this article.
The following is the established format for referencing this article:
Melles, S., S. Glenn, and K. Martin. 2003. Urban bird diversity and landscape complexity: Species–environment associations along a multiscale habitat gradient. Conservation Ecology 7(1): 5. [online] URL: http://www.consecol.org/vol7/iss1/art5/
Report Urban Bird Diversity and Landscape Complexity: Species–environment Associations Along a Multiscale Habitat Gradient Stephanie Melles1, Susan Glenn2, and Kathy Martin3
1University of Toronto; 2Gloucester County College; 3University of British Columbia
- Abstract
- Introduction
- Methods
- Results
- Discussion
- Responses to This Article
- Acknowledgments
- Literature Cited
For birds in urban environments, the configuration of local habitat within the landscape may be as critical as the composition of the local habitat itself. We examined the relative importance of environmental attributes (e.g., tree cover, composition, and number of tree species) measured at different spatial scales in relation to urban bird species richness and abundance. We expected that some bird species and nesting guilds would have a closer association with landscape-level features (within 1000 m), such as proximity to large forested areas, than with local-scale habitat measures (within 50 m). To investigate this, avian community data were collected at 285 point-count stations in 1997 and 1998 along four roadside transects located in Vancouver and Burnaby, British Columbia, Canada. Transects (5–25 km in length) bisected three large parks (>324 ha) and proceeded along residential streets in urban and suburban areas. In total, 48 bird species were observed, including 25 common species. Species richness declined in relation to a gradient of increasing urbanization, as measured by local- and landscape-level habitat features. We further examined the significance and importance of local- vs. landscape-level habitat attributes using logistic regression and found that both scales explained the presence/absence distributions of residential birds. Local-scale habitat features such as large coniferous trees, berry-producing shrubs, and freshwater streams were of particular importance in estimating the likelihood of finding bird species. Landscape measures, particularly forest cover (within 500 m) and park area (measured at different scales as a function of distance from point-count stations) significantly improved likelihood estimations based solely on local-scale habitat features. Our results suggest that both local- and landscape-scale resources were important in determining the distribution of birds in urban areas. Parks, reserves, and the surrounding residential areas should be integrated into urban planning and development designs to maintain resident avifauna and overall species diversity in urban environments.
KEY WORDS: bird response to urbanization, bird species diversity, British Columbia, canonical correspondence analysis, Greater Vancouver, local vs. landscape scales, logistic regression, sequential logistic regression, site occurrence, source habitat, urban habitat gradient.
Published: February 27, 2003
Worldwide, urban areas are expanding both in size and number. As a result of urban expansion, native vegetation is reduced and fragmented over a landscape mosaic in which both the amount of impervious surface is increased, and the structure and composition of the remaining vegetation is progressively altered (Beissinger and Osborne 1982, Arnold and Gibbons 1996, Germain et al. 1998, Marzluff et al. 1998). Cities are typically located near large water bodies, rivers and estuaries, or along coastlines (40% of cities with populations > 0.5 x 106 are located on coasts; World Resources Institute 1996). Thus, a high proportion of urban areas are situated on the productive bottomland and riparian systems that traditionally support rich vertebrate communities (Knopf et al. 1988, Ohmart 1994). As a consequence, large parks and reserves in urban areas may support high species diversity because these protected areas are the habitat “fragments” of highly diverse ecosystems (Schaefer 1994). Increasing urbanization adjacent to natural areas and parks often results in simplified habitats and a community of birds with fewer species dominated by abundant non-native species (Marzluff et al. 1998).
Conservation biologists have focused predominantly on the protection of “natural” ecosystems and have placed little importance on urban areas or urban biodiversity overall (Jules 1997, Vandermeer 1997). A few studies on birds in low-density residential areas (e.g., suburbs that often have a high diversity of habitat types) have shown that these areas may have potential for land management practices enhancing the value of these areas for birds (DeGraaf 1991, Blair 1996). However, vegetation is invariably altered with urbanization. Suburban areas rarely include the full complement of vertical strata found in natural forests (Beissinger and Osborne 1982), and native plant species are often removed or replaced by exotic ornamentals (Rosenberg et al. 1987, Blair 1996). In addition, many urban and suburban areas are located close to undeveloped native areas or forests. As a result, birds may respond directly to local characteristics of the vegetation within urban habitats, but may also respond to broader landscape features including the proximity of large forested areas and developed areas.
The distribution of birds in urban areas has been investigated in relation to at least three dominant ecological theories. In the past, stand-level habitat models compared species assemblages in relation to the structure and composition of (urban) resources at the local or stand level (Emlen 1974, Campbell and Dagg 1976, Lancaster and Rees 1979, Edgar and Kershaw 1994). Under this perspective, the habitat requirements of individual species were examined at the local scale (2–25 ha; Weber 1972, Emlen 1974, Campbell and Dagg 1976, Lancaster and Rees 1979), but the influence of habitats surrounding each stand (city block) was generally not considered. Relating local bird abundance to stand-level habitat factors alone may be more appropriate under conditions where the local habitat quality is high and where nesting success is also high (Arcese et al. 1992, Brawn and Robinson 1996). However, this perspective, if applied under conditions of poor-quality local habitat, neglects to consider the influence of more productive, forested areas nearby (Sewell and Catterall 1998). Further, numerous studies in rural or forested areas have emphasized the importance of landscape-scale structure on the distribution and abundance of many species.
Second, island biogeography theory and metapopulation theory relate island isolation and habitat patch size to species richness, immigration, and extinction rates, and have been applied to habitat patch dynamics in fragmented urban areas (MacArthur and MacArthur 1961, MacArthur and Wilson 1967, Tilghman 1987, Soulé et al. 1988, Hanski 1999). Some studies of birds in urban areas have focused on species richness within a patch of native habitat (e.g., a large park) in relation to patch size and the degree of isolation from other areas of habitat (Tilghman 1987, Diamond 1988, Soulé et al. 1988). Soulé et al. (1988) related the absence of Spotted Towhee (Pipilo maculatus) to its inability to disperse to and survive in small habitat patches within a presumed uninhabitable sea, but this study largely ignored other habitat features in the area. The absence of towhees may have been more related to the absence of local and landscape habitat attributes than to the birds’ inability to disperse to or survive in these small habitat patches. Research relating local species occurrence and diversity patterns to island biogeography or metapopulation theory may apply when the landscape can be classified into suitable and unsuitable habitat, as with islands vs. ocean. Urban habitats are altered from their original state, but do offer foraging and cover resources and are thus not always hostile or unsuitable for all species. In a complex landscape mosaic, the presence and detection of individual birds is expected to vary with land use and with the overall composition of the landscape (Trzcinski et al. 1999, Austen et al. 2001, Fahrig 2001). The focus on isolation of strict habitat patches is not sufficient in many scenarios.
Thirdly, the distribution of birds in urban areas has been investigated using spatial-structuring theories of ecological communities in which local-scale habitat features are viewed in relation to the surrounding landscape features (e.g., Bolger et al. 1997, Rottenborn 1999, Saab 1999, Odell and Knight 2001). Spatial-structuring studies have often indicated that the surrounding landscape characteristics (100 ha to <30,000 ha) provide significant additional information to local-scale bird habitat models that include only local, site-level (<3 ha) habitat characteristics (Smith and Schaefer 1992, Blair 1996, Bolger et al. 1997, Germain et al. 1998, Rottenborn 1999, Mörtberg 2001). However, the ecological mechanisms generating community responses to landscape-level spatial variability have not been well established (Bolger et al. 1997, Germain et al. 1998, Marzluff et al. 1998). Earlier studies acknowledged landscape-level effects (e.g., Emlen 1974, Vale and Vale 1976, Lancaster and Rees 1979), but the complexity of the urban mosaic itself was difficult to characterize. We know of only a few studies that have examined urban bird communities citywide at different spatial scales (Haddidian et al. 1997, Reynaud and Thioulouse 2000). Thus, we chose the spatial-structuring approach to examine avian community composition in relation to features of the entire urban matrix.
Urban areas generally have an intensively developed core surrounded by irregular rings of diminishing levels of development; the environmental variation ostensibly is ordered in space along some kind of gradient. Urban gradient analysis attempts to capture this gradient of environmental variation and to examine the response of the biotic community (McDonnell and Pickett 1990). Although a few studies have examined the effects of surrounding urbanization on bird species richness in park and riparian fragments (e.g., Saab 1999, Sodhi et al. 1999, Mörtberg 2001, Er 2002), none has looked at the reverse, that is, the effects of surrounding parks on the bird communities in residential areas. The occupancy of poor-quality or marginal areas by resident birds could be influenced by the surrounding landscape (regional parks vs. high-density housing) because the surrounding habitats may act as resource areas for residential birds and as “source” areas for dispersing birds.
To assess the relative importance of local- and landscape-level habitat measures in relation to observed bird distributions, we used urban gradient analysis to examine the bird community in Vancouver and Burnaby, British Columbia, Canada along four transects. Specifically, we tested the expectation that bird species richness should decline and mean relative abundance of the remaining species should increase with increasing urbanization, as summarized by a habitat gradient. We expected that the surrounding landscape (habitat measures within 0–1000 m) would adequately describe this urbanization gradient and would make better predictors of bird species and nesting guild presence than local-level habitat measures at the residential plot scale (within 50 m). We were also interested in examining whether or not species incidence (the proportion of sites occupied) increased with park proximity, possibly because birds disperse from high-density park areas or because parks contribute critical resources to nearby marginal residential areas.
The municipalities of Vancouver and Burnaby, hereafter "Greater Vancouver," British Columbia (49°18' N, 123°12' W; Fig. 1), are located within the Georgia Basin ecoprovince of British Columbia. The Georgia Basin represents only 3% of the British Columbia land base (2,772,571 ha), yet supports the highest diversity of breeding birds in the province, contains three cities, and approximately three-quarters of the provincial population, 2.9 x 106 people on the Canadian side (Demarchi 1996). Greater Vancouver contains several relatively large parks distributed unevenly over the urban mosaic: Pacific Spirit, Stanley, Central Park, and Vancouver and Burnaby Regional Parks (Fig. 1). These parks still contain representative vegetation of the Coastal Western Hemlock zone (Appendix 1) and also contain many species of non-native vegetation, such as English holly (Ilex aquifolium) and Himalayan blackberry (Rubus discolor). Please refer to Appendix 1 for further details on the study area.
Bird surveys
We collected data on the relative abundance of individual bird species at 285 point-count locations along four roadside transects in Vancouver and Burnaby (Fig. 1). Point locations (with an interstation distance of 250 m) were sampled only once in each year during the breeding season, 24 June–13 July 1997 and 1 May–1 June 1998, to maximize the number of sites that could be surveyed over the landscape. Birds flying over the station were not recorded because they were unlikely to be breeding in the area. We used a 50-m fixed-radius methodology and a count duration of 5 min (DeGraaf et al. 1991, Ralph et al. 1993). All bird surveys were conducted by the primary author on clear days during the first 4 h following sunrise, to coincide with peak singing activity.
Point-count data were collected only once per breeding season over each of two years, and the timing of data collection was much later in 1997 than in 1998. This may limit the inferences that we can make about population and community trends. However, in order to investigate landscape-level bird–habitat trends, it is necessary to maximize the number of sites that can be surveyed over the landscape in any one season (Austen et al. 2001). Replication in space rather than performing multiple point counts at fewer locations can be an advantage rather than a disadvantage, because replication in space leads to more certainty about the species–habitat associations at the expense of certainty about a particular species’ presence at any individual point (Bolger et al. 1997, Goodinson 2000). Bird names follow Campbell (1998).
Habitat characteristics: local variablesCity zoning base maps (1:2,000) were used to delineate the 50-m boundaries for vegetation plots in order to estimate local measures of potential bird nesting habitat (Table 1). The percent cover of grass (to a maximum of 100%) was estimated by sketching grass cover onto these zoning maps for each residential lot within the boundary of the circumscribed 50-m plots. These maps were later overlain with dot-matrix acetate to estimate local percent grass cover (LOCGRASS). We found it more expedient, however, to simply count the number of house lots (LOTS) than to estimate impervious surface cover in an analogous way: sketching houses, garages, driveways, sidewalks, and streets onto city zoning base maps and overlaying again with dot-matrix acetate was too time consuming. Moreover, research has shown that housing density is a good estimate of impervious surface cover (Dinicola 1990). The number of house lots was adjusted in commercial and industrial areas because single buildings often covered the entire lot in these areas. When this was the case, we increased the number of house lots by 15 and by 10 if apartment buildings were present, because the average number of house lots in multi-family areas was nine and we wished to have a comparable estimate for the number of house lots across all sites. Elevation (ELEV) was estimated using TRIM (Terrain Resource Information Management) maps (1:20,000) with contours generated from a digital elevation model (TRIM 1993).
Landscape variables
Landscape cover variables were measured from digital aerial photographs (May/July 1995, 1:30,000) with an orthophoto accuracy registered to TRIM (Terrain Resource Information Management, 1:20,000) and a pixel size of 1 m. An acetate dot-grid overlay, representing a 500 m radius circle, was centered on each survey site located on the aerial photographs and percent cover values were recorded for grass (GRASS500), impervious surface (IMPERV500), coniferous trees (CONIF500), and deciduous trees (DECID500); see Blair (1996), Bolger et al. (1997), Germaine et al. (1998), and Rottenborn (1999).
A digital map of land use in the Greater Vancouver Regional District (GVRD) was used to develop a map of green space in the study area (GVRD 1996), including parks > 1 ha, cemeteries, and golf courses (IDRISI version 4.0, GIS software; Eastman [1992]). Universal Transverse Mercator (UTM) coordinates for the point-count stations were registered to the map of urban green space. We derived four distance–park landscape metrics at increasing spatial scales (0–260 m, 0–500 m, 0–760 m, and 0–1000 m) to summarize the amount and proximity of park area around each station (PARK260, PARK500, PARK760, PARK1000).
The four park area by distance metrics were calculated by dividing the proportion of park area (within 20-m concentric rings around each bird counting station) by distance from the point-count station and summarizing over all concentric rings included in each scale (0–260 m, 0–500 m, 0–760 m, and 0–1000 m, cumulatively increasing), as follows (Eq. 1): P = park area within each 20-m ring, and D = distance (i.e., 20 m, 40 m, 60 m...), summed over k distance rings within each park area by distance variable. For example, for the park index 0–260 m, the total radius, 260 m, is divided by 20-m rings (260/20 = 13 rings, k = 13). Dividing by distance effectively downweights the park area by the distance from survey locations.
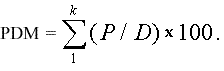 |
(1) |
Species that were observed only once (12 species) were removed from the analyses to avoid potentially mistaking migrants or wandering individuals for residents; species observed on <5% of the sites (8 species) were made supplementary, having no influence on further analyses, and are presented in the canonical correspondence analysis (CCA) ordination diagram only. Moreover, three species of gulls were excluded from analyses, for a total of 23 excluded species. Because birds were surveyed later in the 1997 breeding season (24 June–13 July) than in the 1998 breeding season (1 May–1 June), correlation analysis was used to test for year effects by examining the relationships between the relative abundance of different nesting guilds by year. We did not attempt to account for repeated measures by year, and found no significant year effects. Thus, data were pooled for later analyses (Melles 2001). The landscape metrics were not independent from point to point, so we reduced the data set by a factor of four for statistical analyses to maintain independent landscape metrics for each response term. Species data, local-, and landscape-level environmental variables were averaged over each of four points (minimum 4 x 250 m = 1000 m apart), reducing the sample size to 70 points. The data were tested for normality, skewness, and kurtosis to determine the need for data transformations (ter Braak 1986, Hosmer and Lemeshow 1989, Jongman et al. 1995, Tabachnick and Fidell 1996).
To examine the hypothesis that environmental variability in urban areas is ordered in space along a gradient that results in a simplified bird community with fewer, more abundant species, we used canonical correspondence analysis (CCA). CCA is akin to direct gradient or regression analysis done in multivariate space. The species matrix consisted of the maximum abundance distributions of 25 species of birds; this matrix was related to a linear combination of 29 local- and landscape-level habitat variables (Table 1). For the CCA, the highest (maximum) relative abundance recorded in 1997 or 1998 was used instead of an average, to avoid the smoothing effect of an average that would result in less detectable bird–habitat trends. Although the maximum value may be an optimistic estimate, this measure is likely to be a more accurate estimate of abundance at a particular site than the mean of one survey in each of two years (Vander Haegen et al. 2000). The significance of the CCA ordination of species–habitat relationships was investigated by performing a randomization test (Monte Carlo) on the projected relationships; 199 random permutations were performed on the significance of the ordination axes (CANOCO 4; ter Braak and Šmilauer 1998). With canonical correspondence analysis, one can infer which variables, local and/or landscape, may best explain the variation in species distributions, because the most important habitat variables load most highly on the first axis.
Species richness as opposed to species abundance was expected to decline with increasing urbanization, as summarized by a habitat gradient; therefore, we graphed and performed simple regression on the total number of species (including all of the uncommon species) vs. the first axis of the composite urban gradient. Using logistic regression, we tested the importance of variables at increasing spatial scales in estimating the likelihood of detecting different nesting guilds and individual species. In addition, to explore directly the relationship between individual species incidence (i.e., the proportion of stations occupied) and the amount of park area in close proximity, we plotted increasing park area by distance metrics vs. the proportion of sites occupied, and used simple regression to examine the shape of this relationship.
Subsets of species, particularly ground- and shrub-nesting bird guilds, are of concern to land managers in urban areas because residential zones may be currently unsuitable for these groups (Rottenborn 1999). Birds were grouped into the following nesting-habitat guilds: deciduous tree, coniferous tree, building, ground, shrub, and cavity nesters, as per Ehrlich et al. (1988), with three exceptions. The Violet-green Swallow (Tachycineta thalassina) and the European Starling (Sturnus vulgaris) were assigned to the building-nesting guild, because in urban areas they more commonly nest in buildings. The Brown-headed Cowbird (Molothrus ater) was assigned to the ground-nesting guild because we decided that these nest parasites should be grouped with their most common host species, the Song Sparrow (Melospiza melodia). Grouping birds according to their nesting guild should provide useful information about habitat restrictions and should allow for more powerful statistical tests than when considering each species separately.
However, when the data set was reduced by a factor of four, only two guilds had distributions that were not present at all 70 sites: the ground and shrub nesters. Thus, representative species of each guild were selected for pursuant regression models when sample sizes for that representative species were sufficient to create a robust model. Six species (Bushtit (Psaltriparus minimus), Pine Siskin (Carduelis pinus), Violet-green Swallow (Tachycineta thalassina), Spotted Towhee (Pipilo maculatus), Song Sparrow (Melospiza melodia), and Winter Wren (Troglodytes troglodytes) were selected to represent the different guilds. There were no suitable representative species for the shrub nesters, but this guild was modeled as a whole. Two species of ground-nesting birds were selected for the modeling process because species in this guild tend to be highly selective and we wanted to investigate the differences between relationships detected for the guild as a whole and relationships detected on a species-by-species basis.
The significance of the two spatial scales of variables was tested using sequential logistic regression. With sequential logistic or “block” regression, terms were entered into a model in separate blocks wherein all local terms represented one block and all landscape terms represented another. For each model group, we fit either the local or landscape blocks of variables first, and then determined whether or not landscape-level variables improve the prediction of bird presence beyond that of local variables alone, or the reverse: landscape followed by local blocks. We used stepwise logistic regression (Pforward <0.20, Pbackward <0.25; Hosmer and Lemeshow 1989, Tabachnick and Fidell 1996) to statistically select which variables we should enter at each block, for individual species and guilds. Only a subset of the original 29 local and landscape variables were entered in the sequential block models (generally only two to four local variables and one to two landscape-level variables were entered at each block). The final models were evaluated according to their biological relevance and statistical significance. All logistic regression analyses were done using SPSS (1996) and model residuals were examined for deviations from the assumption of linearity in the logit.
All four of the park area by distance indices were highly correlated (Pearson’s r >0.9), which is not surprising, as they contain overlapping information. We were interested in examining the effect of these indices at increasing scales; although larger scales included the same information as smaller scales, birds presumably respond to their environment at cumulative distances from their surroundings, rather than responding to “donuts” of habitat located 500–760 m away, for example. Impervious surface cover was highly correlated with all of the park area by distance indices and with coniferous/deciduous tree coverage (Pearson’s r = -0.68 to -0.81). Coniferous and deciduous tree cover was correlated with park area by distance indices as well (Pearson’s r = 0.41–0.60). Most of the landscape-level variables were not as highly correlated with local-level variables (Pearson’s r < 0.55), indicating generally no more than 30% shared variation between sets of paired local- and landscape-level variables. With the exception of the house lots variable, which was negatively correlated with park aea by distance indices (Pearson’s r~ -0.65) and was positively correlated with impervious surface cover (Pearson’s r = 0.81), and GRASS500 cover, which was correlated with LOCGRASS (Pearson’s r < 0.63), the majority of the local variables were not highly correlated with each other (Pearson’s r <0.4).
Forty-eight bird species were observed, including 25 common species that were retained for the analyses. Two urban non-native species were typical on almost all sites: the ubiquitous European Starling (Sturnus vulgaris) and House Sparrow (Passer domesticus); non-native species had the highest maximum relative abundance on all occupied sites (Table 2). Thus, like most other urban habitats, Greater Vancouver contains a greater proportion of exotic birds than native bird species (Case 1996). The most common species, in terms of the number of sites detected, were European Starling (Sturnus vulgaris), Northwestern Crow (Corvus caurinus), House Sparrow (Passer domesticus), House Finch (Carpodacus mexicanus), American Robin (Turdus migratorius), and Black-capped Chickadee (Poecile atricapillus; Table 2). Building nesters had the highest relative (maximum) abundance per occupied site as they were detected up to three times more often than other guilds. Although urban habitat was the dominant habitat type on most sites, building nesters were more abundant than other bird guilds in their constituent habitats, indicating that building-nester abundance was disproportionately high. In sequence, the deciduous tree and coniferous tree nesters followed building nesters in the number of occupied sites and the relative (maximum) abundance per occupied site. Moderate numbers of cavity, ground, and shrub nesters were detected per site occupied (Table 2).
Bird community–habitat relationships, CCA
The relationships between community composition and habitat variables are depicted by a joint plot of bird species scores (points) in relation to habitat variables (arrows, CCA ordination; Fig. 2). Both the length of the habitat arrow on ordination diagrams and the correlation coefficients for the variable provide an indication of relative importance for that variable (Table 1). Because some variables contained overlapping information with increasing scale (i.e., the park area by distance metrics), these variables were highly correlated. Following ter Braak and Šmilauer (1998), we removed variables with variance inflation factors above 20 from the CCA. However, the correlation of habitat variables with the axes do not become unstable when variables are confounded. Although the CCA changed when these variables were removed, the correlation of these variables with the axes did not change, and thus they were also presented in Table 1 (ter Braak and Šmilauer 1998). Supplementary environmental variables are useful to provide an alternative interpretation of the ordination diagram, but they do not influence the definition of the ordination axes (ter Braak and Šmilauer 1998). The degree of correlation with Axis I increased with increasing measurement scales from 260 m to 1000 m radii for all of the overlapping park area by distance metrics (Table 1). Impervious surface cover (IMPERV500) had the highest correlation with Axis I overall (-0.81), and number of house lots (LOTS) had the highest correlation with Axis I (-0.61) among the local-level variables.
Despite the demonstration of an urbanization gradient in this data set, much of the spatial variation in the habitat did not follow the expected, gradual decline from a highly developed urban core to more natural areas. In fact, the four roadside transects sampled follow a rather irregular gradient traversing a complex mosaic including neighborhoods with a few large parks and several medium and small parks (Fig. 3). All four transects progressed through and away from one large park, but three came close to at least one other moderate- or large-sized park (Fig. 3). Generally, the effect of these different parks on the variables impervious surface cover (IMPERV500) and number of house lots (LOTS) emulated the variable PARK1000, except that where PARK1000 had high values, impervious surface cover and the number of house lots had low values (Fig. 3). The variability in tree species richness was less patterned because many urban and suburban areas plant a diversity of non-native tree species, but this local-level variable tended to vary inversely with LOTS.
The CCA ordination shows how bird species respond to several key environmental variables (both local and landscape) along a gradient of urban mosaic complexity (Fig. 2). Axis I can be viewed as an intensity of urbanization gradient characterized largely by variables such as deciduous and coniferous tree cover, park area by distance indice(s), and dead and downed wood at the most natural end. Impervious surface, housing density, and intersection size characterized the most urban, negative end of Axis I. Thus, the two ends represent extremes in the urban mosaic. At one extreme, regional parks contain large amounts of natural habitat, and at the other extreme, heavily developed areas contain more urban structure. Axis II (vertical) can be viewed more as a diversity of local vegetation axis characterized predominantly by large coniferous shrubs, tree species richness, and large coniferous trees. The overall ordination is significant, indicating that the ordination provided a reasonable explanation of the relationships between observed bird species distributions and the two scales of habitat variables, local and landscape (P<0.01, Monte Carlo randomization test; CANOCO 1998).
In total, 26 of the 33 species (including eight “rare” supplementary ones) were located on the natural end of the urbanization gradient, including seven of eight ground-nesting species, all five shrub-nesting species, and four cavity-nesting species. Because this side of Axis I was dominated by landscape variables, there was some support for the expectation that landscape variables might adequately describe the urbanization gradient (Fig. 2, Table 1). When we examined the relationship between species richness and the urbanization gradient, there appeared to be a potential threshold below which species richness declined more quickly (Fig. 4; curvilinear regression with two significant terms, P<0.05).
Evaluating local- and landscape-level effects on bird–habitat relationships
Six species were selected to evaluate the influence of local- and landscape-scale variables. When variables at both scales were allowed to enter in stepwise logistic screening models, local variables were represented as the first variable entered for four of six bird species. Once we accounted for the local habitat, landscape-scale variables explained significantly more variation in the distribution of four of the six species (Tables 3 and 4). Thus, only two species had no association with features at the landscape level: Violet-green Swallows and Bushtits. These species were more associated with local variables such as the numbers of coniferous trees, the presence of fresh water (STREAMS), and housing density (LOTS), rather than measures of landscape cover (Table 4). Swallows in particular seem well adapted to the urban environment in Greater Vancouver and were detected across the entire gradient. Conversely, Winter Wrens and shrub-nesting species were more likely to be detected within parks or areas with large parks in close proximity (as measured by PARK760 and PARK1000; Table 4). Thus, when landscape effects were entered first into these models, the local-level habitat did not significantly improve estimates of the occurrence patterns of wrens and shrub nesters (Table 3).
For four of the six individual species that were modeled, local habitat variables alone were more precise in terms of likelihood ratio tests than were landscape variables alone. Local-level habitat variables sometimes lost or declined in significance (P<0.2) when landscape-level variables were added to the models, but we retained these variables if they were considered biologically significant. Several species, including ground-nesting species, were at least three times more likely to be detected near water, although only towhees were significantly so (i.e., streams odds ratio; Table 4), highlighting the possible importance of water for a variety of bird species. The odds ratio is simply the exponent of variable coefficients and represents the increase in the odds of detecting different bird species per unit increase in the related variable. For every 50-m residential plot with a minimum of nine mid-sized conifers, Winter Wrens were almost four times more likely to be present, Bushtits were 11 times more likely, and Pine Siskins were 17 times more likely to be present (exponent of coefficients; Table 4). The effects of large berry-producing shrubs (i.e., salmonberry, blackberry, and elderberry) were similar for Spotted Towhees and Song Sparrows (Table 4), indicating that variation in the local bird habitat could have a substantial influence on species occurrence. In general, the coefficients for significant effects of landscape-level variables were smaller than local variable coefficients (Table 4). The increases in odds for every 10-unit increase in the index, PARK760 or PARK1000, were smaller than local variable odds ratios and ranged between a 10% and a 20% increase.
Bird species incidence as a function of park area by distance indexThe occupancy of stations with similar habitat characteristics should increase both with area and proximity to parks if parks are source areas for marginal nesting locations along residential streets. The relationship between the four park area by distance metrics at increasing scales and selected bird species or nesting guilds were investigated using simple regression; only the most significant relationships were graphed in Fig. 5A–D. Species were selected if they were present on more than 10 sites, if less than two-thirds of all sites were occupied (i.e., they were not ubiquitous), and if they were significantly related to landscape-level habitat variables in the simple regression models including only these landscape variables. In all cases, the largest scale habitat metric, PARK1000, explained the greatest amount of variation in the occurrence patterns of American Robin, Spotted Towhee (45% and 40% variation explained), and ground- and shrub-nesting guilds (43% and 57%, respectively). Thus, regional park space was an important factor influencing the presence of these species and groups of birds (Fig. 5A–D; P<0.001). Still, it is important to reiterate that local housing density (LOTS) correlated negatively with these park area by distance indices and could also account for the shape of this relationship. The shape and extent of the influence of park area by distance indices also varied by species, with some species being more sensitive to large, proximal park areas. The variability around the relationship is probably related to features of local-level habitat.
The global trend of increasing urbanization has been shown repeatedly to result in depauperate bird communities. Our results were consistent with other studies and results from other cities showing that the number of species declines with increasing urbanization and that the remaining group of species is dominated by highly abundant species (e.g., Emlen 1974, Campbell and Dagg 1976, Lancaster and Rees 1979, Edgar and Kershaw 1994). The patterns of urbanization, however, have rarely been examined in terms of the habitat mosaic, including both local- and landscape-level attributes characterized by gradient analysis (but see Blair 1996, Bolger et al. 1997, Rottenborn 1999, Reynaud and Thioulouse 2000). In the present study, when we characterized the gradient to include mosaic composition, interesting community trends emerged. Many sensitive groups of species (e.g., ground and shrub nesters) were associated with both local- and landscape-level habitat features including streams, deciduous trees, and high amounts of surrounding parks and forest cover. With reduced forest cover, the species composition changed and the community became dominated by more “urban-adapted” species such as Black-capped Chickadees, Bushtits, Violet-green Swallows, American Robins, and House Finches, with distributions that were related to high amounts of local-level habitat (i.e., coniferous and deciduous trees and shrubs; Fig. 2). Increasing impervious surface cover, combined with fewer trees, resulted in a community dominated by four to five “urban” bird species, three of which were non-native species (House Sparrows, European Starlings, and Rock Doves; Fig. 2).
Local vs. landscape habitat predictorsThe evidence to support the hypothesis that broad-scale habitat measures are better predictors of bird species and nesting guild presence than local-level habitat measures was not conclusive, partly because habitat relationships at the two scales were confounded. Landscape forest cover and park area by distance metrics did, however, add significantly to model predictions for four of six species and two nesting guilds, once specific features of the local habitat were included. The reverse was also equally true, as local-level variables added significantly to the same number of models when we accounted for parks and surrounding forest cover. Other studies have demonstrated support for landscape-level effects on birds in urban areas and generally have concluded that urbanization around forest patches and riparian areas affects bird diversity and abundance within these areas (e.g., Munyenyembe et al. 1989, Smith and Schaefer 1992, Bolger et al. 1997, Germaine et al. 1998, Saab 1999). However, results from multiple-scale studies have not consistently shown that these effects are significant (Berry and Bock 1998, Clergeau et al. 1998). Our results suggest that urban bird communities were “rescued” by their surroundings because several species were associated with landscape forest cover and parks along the urban gradient.
Fresh water (streams), large conifers (30–60+ cm dbh), and large berry-producing shrubs such as salmonberry, blackberry, and elderberry were local variables that consistently showed significant relationships with the presence of a variety of bird species. Some species could be classified as sensitive species (i.e., Spotted Towhees and Song Sparrows) and were detected only in areas with particular local habitat site characteristics (fresh water and shrubs) within regions that also had high amounts of forest and park cover. At the landscape scale, the most important predictors were park area by distance metrics, PARK760 and PARK1000 (Table 4), so neighborhoods immediately surrounding large parks had a greater proportion of different bird species than did highly urban areas. Neighborhoods surrounding parks also typically had larger house lots (recall that areas with high park area by distance indices also had fewer house lots; hence, this negative correlation was possibly related to unmeasured socioeconomic factors). These areas might increase the likelihood of attracting sensitive species by improving features of the local habitat (see Conservation implications). For shrub- and ground-nesting guilds as well as Spotted Towhees and Song Sparrows, landscape variables alone were more precise predictors than local variables, illustrating the diversity in response by birds to both habitat structure and spatial scale. Given that there were one to two more descriptors of the local habitat compared to landscape level descriptors, local blocks may include more biological information, although it could be argued that each landscape measure summarized more biological information by representing a larger spatial scale.
Bird species incidence as a function of park area and distanceOur park area by distance metrics represent the proportion of park area within concentric rings around each site, divided by distance from the station. We thus downweighted park area by the inverse of distance to account for the fact that the outermost rings were much larger than the innermost ring and to investigate the effect of distance from parks on birds. This inverse of distance weighting scheme assigned a decreasing, nonlinear weight to park area as a function of distance and was related to the theoretical expectation that species and guild occurrence should decrease nonlinearly with distance from “natural” areas, given the dispersal distance trends for several species of birds examined by Sutherland et al. (2000). We found significant relationships between park area by distance metrics and the distributions of American Robins, Spotted Towhees, and shrub- and ground-nesting species, but our results were not consistent with those of Bolger et al. (1997), who created an urban exposure index by assigning an inverse weighting scheme to the proportion of developed land within each concentric ring around point counts located in “islands” of natural habitat, creating an urban exposure index. When they entered this variable in logistic regression models, urban exposure was significant in only two of 20 species occurrence habitat models. Bolger et al. (1997) found more significant variables to be the nonweighted proportions of edge and natural areas around “island” survey sites.
Conservation implicationsCurrently, only 5% of the lower mainland of British Columbia, including Greater Vancouver, is set aside as parkland (80,000 ha of 1.58 x 106 ha; GVRD 1993, 1996). Although a large portion of the area maintains some natural character under forest management and agricultural reserves, by the year 2021, more than 1.2 x 106 additional people are expected to live in the Greater Vancouver area. Our results suggest that multiple elements of the landscape mosaic at both local and landscape scales are important in determining the distribution of birds in urban areas, so parks and areas surrounding parks and reserves should be integrated into urban planning and development designs if avian diversity is to be considered in conservation or management actions. Development on the verge of continuously forested areas should minimize impervious surface cover, minimize house size, maintain native tree cover and berry shrubs, integrate new ponds, and maintain and develop natural freshwater sources into planning designs. Residential areas near parks have a high likelihood of recruiting sensitive nesting species and will probably experience frequent use by species from nearby parks.
Speculation: natal dispersal, landscape complementation, and supplementationNumerous processes could be involved in the patterns that we observed here. For instance, the dispersal of birds from larger forests in the region may determine the bird assemblages of other smaller forest “patches” (Freemark and Merriam 1986, Hinsley et al. 1996). Dispersing individuals may be more likely to settle near their natal area or near conspecifics, which may increase their chances of finding or attracting a mate (Austen et al. 2001). Species may supplement resources in small urban habitat patches with nearby resource patches in large parks, or species may complement resources (such as bird feeders) in surrounding residential areas with nesting locations inside the park (landscape complementation and supplementation; Dunning et al. 1992, Er 2002). Moreover, birds may be unwilling to cross large gaps, so connectivity becomes an issue in highly fragmented urban areas (Desrochers and Hannon 1997).
If the spatial patterns of birds that we observed are generated by dispersal and settlement biases (Bolger et al. 1997), then landscape habitat characteristics such as proximity of habitat to natal “source” areas will affect the chances that a bird species will occur at a particular site. Habitat specialists are likely to disperse from reserve areas in times of high regional nesting success to more marginal nesting areas, where they may be exposed to increased predation and competition (Soulé et al. 1988, Hinsley et al. 1996, Helzer and Jelinski 1999, Rottenborn 1999, Burke and Nol 2000). Thus, further research on bird dispersal and settlement patterns may help to explain the mechanism of spatial structuring in bird communities, especially in urban areas. Some evidence suggests that birds search for and select breeding territories for the following year during their postfledgling exploration (Baker 1993). Therefore, habitat attributes and proximity to the natal area will affect the occupancy of areas close to successful breeding sites (Baker 1993, Bolger et al. 1997, Helzer and Jelinski 1999, Sutherland et al. 2000).
Although landscape-level studies detect and predict patterns, local-level and demographic studies are necessary to determine many of the mechanisms involved in avian population and community change. Urban bird studies should examine a few key landscape measures, such as distance to and size of large natural park areas with habitat offering natural conditions for birds and the processes that are affected by these variables (e.g., Er 2002). We also recommend that impervious surface be measured at different spatial scales to facilitate comparison with different cities. Overall, such multiscale examinations of attributes to characterize avian habitat associations will improve our understanding of urban ecology and will allow us to determine the optimal balance of intensive local-level and extensive landscape-level studies needed to understand both the processes and patterns that maintain or restore a high proportion of native urban bird communities.
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a comment, follow this link. To read comments already accepted, follow this link.
This project was funded, in part, with generous support from the Asa-Yohal Graduate Fellowship in Forestry and the Bert Hoffmeister Scholarship in Forest Wildlife. The Centre for Applied Conservation Biology, the Geography Department at University of British Columbia, and the Canadian Wildlife Service also provided many resources and support. The authors gratefully acknowledge the comments of two anonymous reviewers whose suggestions substantially improved the manuscript. Dr. Brian Klinkenberg, Dr. Jamie Smith, and Dr. Marie-Josée Fortin gave their expertise, comments, and assistance generously and we gratefully acknowledge their reviews and support as well.
Arcese, P., J. N. M. Smith, W. M. Hochachka, C. M. Rogers, and D. Ludwig. 1992. Stability regulation and the determination of abundance in an insular Song Sparrow population. Ecology 73:805–822.
Arnold, C. L., and C. J. Gibbons. 1996. Impervious surface cover: the emergence of a key environmental indicator. Journal of the American Planning Association 62:243–255.
Austen, M. J. W., C. M. Francis, D. M. Burke, and M. S. W. Bradstreet. 2001. Landscape context and fragmentation effects on forest birds in southern Ontario. Condor 103:701–714.
Baker, R. R. 1993. The function of post-fledging exploration: a pilot study of three species of passerines ringed in Britain. Ornis Scandinavica 24:71–79.
Beissinger, S. R., and D. R. Osborne. 1982. Effects of urbanization on avian community organization. Condor 84:75–83.
Berry, M. E., and C. E. Bock. 1998. Effects of habitat and landscape characteristics on avian breeding distributions in Colorado foothills shrub. Southwestern Naturalist 43:453–461.
Blair, R. B. 1996. Land use and avian species diversity along an urban gradient. Ecological Applications 6:506–519.
Bolger, D. T., T. A. Scott, and J. T. Rotenberry. 1997. Breeding bird abundance in an urbanizing landscape in coastal southern California. Conservation Biology 11:406–421.
Brawn, J. D., and S. K. Robinson. 1996. Source–sink population dynamics may complicate the interpretation of long-term census data. Ecology 77:3–12.
Burke, D. M., and E. Nol. 2000. Landscape and fragment size effects on reproductive success of forest-breeding birds in Ontario. Ecological Applications 10:1749–1761.
Campbell, C. A., and A. I. Dagg. 1976. Bird populations in downtown and suburban Kitchener-Waterloo, Ontario. Ontario Field Biologist 30:1–22.
Campbell, R. W. 1998. British Columbia birds: a species list. Common and scientific names, sequence, and 4-letter codes. Wildlife Report Number 2, Wild Bird Trust of British Columbia, West Vancouver, British Columbia, Canada.
Clergeau, P., J.-P. L. Savard, G. Mennechez, and G. Falardeau. 1998. Bird abundance and diversity along an urban–rural gradient: a comparative study between two cities on different continents. Condor 100:413–425.
DeGraaf, R. M. 1991. Winter foraging guild structure and habitat associations in suburban bird communities. Landscape and Urban Planning 21:173–180.
DeGraaf, R. M., A. D. Geis, and P. A. Healy. 1991. Bird population and habitat surveys in urban areas. Landscape and Urban Planning 21:181–188.
Demarchi, D. A. 1996. An introduction to the ecoregions of British Columbia. Wildlife Branch, Ministry of Environment, Lands and Parks, Victoria, British Columbia, Canada.
Desrochers, A., and S. J. Hannon 1997. Gap crossing decisions by dispersing forest songbirds. Conservation Biology 11:1204–1210.
Diamond, J. M. 1988. Urban extinction of birds. Nature 333:393–394.
Dinicola, R. S. 1990. Characterization and simulation of rainfall runoff relations for headwater basins in Western King and Snohomish Counties, Washington. U.S. Geological Survey. Water Resources Investigations Report 89-4052.
Dunning, J. B., B. J. Danielson, and H. R. Pulliam. 1992. Ecological processes that affect populations in complex landscapes. Oikos 65:169–174.
Eastman, J. R. 1992. IDRISI. User’s guide. Version 4. Clark University, Worcester, Masachusetts, USA.
Edgar, D. R., and G. P. Kershaw. 1994. The density and diversity of the bird populations in three residential communities in Edmonton, Alberta. Canadian Field-Naturalist 108:156–161.
Ehrlich, P. R., D. S. Dobskin, and D. Wheye. 1988. The birders handbook: a field guide to the natural history of North American birds. Simon and Schuster, Toronto, Canada.
Emlen, J. T. 1974. An urban bird community in Tucson, Arizona: derivation, structure, regulation. Condor 76:184–197.
Er, K. B. H. 2002. Effects of forest loss and fragmentation with urbanization on bird communities in Vancouver. Thesis. University of British Columbia, Vancouver, British Columbia, Canada.
Fahrig, L. 2001. How much habitat is enough? Biological Conservation 100:65–74.
Freemark, K. E., and H. G. Merriam. 1986. Importance of area and habitat heterogeneity to bird assemblages in temperate forest fragments. Biological Conservation 36:115–141.
Germaine, S. S., S. S. Rosenstock, R. E. Schweinsburg, and W. S. Richarsdson. 1998. Relationships among breeding birds, habitat, and residential development in Greater Tucson, Arizona. Ecological Applications 8:680–691.
Goodinson, C. 2000. Limitations of the point count method for inferring stand-level species resource relationships: a sampling simulation approach. Thesis. University of British Columbia, Vancouver, British Columbia, Canada.
Green, R. N., and K. Klinka. 1994. A field guide to site identification and interpretation for the Vancouver Forest Region. Land Management Handbook 28, British Columbia Ministry of Forests, Victoria, British Columbia, Canada.
GVRD (Greater Vancouver Regional District). 1993. Creating Greater Vancouver’s Green Zone. The Livable Region Strategic Plan. Strategic Planning Department, Parks Department, Vancouver, British Columbia, Canada.
GVRD (Greater Vancouver Regional District). 1996. Creating our future. Steps to a more livable region. Strategic Planning Department, Parks Department, Vancouver, British Columbia, Canada.
Haddidian, J., J. Sauer, C. Swarth, P. Handly, S. Droege, C. Williams, J. Huff, and G. Didden. 1997. A city-wide breeding bird survey for Washington, D.C. Urban Ecosystems 1:87–102.
Hanski, I. 1999. Metapopulation ecology. Oxford University Press, Oxford, UK.
Helzer, C. J., and D. E. Jelinski. 1999. The relative importance of patch area and perimeter–area ratio to grassland breeding birds. Ecological Applications 9:1448–1458.
Hinsley, S. A., P. E. Bellamy, I. Newton, and T. H. Sparks. 1996. Influences of population size and woodland area on bird species distributions in small woods. Oecologia 105:100–106.
Hosmer, D. W., and S. Lemeshow. 1989. Applied logistic regression. John Wiley, New York, New York, USA.
Jongman, R. H. G., C. J. F. ter Braak, and O. F. R. van Tongeren. 1995. Data analysis in community and landscape ecology. Cambridge University Press, Cambridge, UK.
Jules, E. S. 1997. Dangers in dividing conservation biology and agroecology. Conservation Biology 11:1272–1273.
Knopf, F. L., R. R. Johnson, T. Rich, F. B. Samson, and R. C. Szaro. 1988. Conservation of riparian ecosystems in the United States. Wilson Bulletin 100:272–284.
Lancaster, R. K., and W. E. Rees. 1979. Bird communities and the structure of urban habitats. Canadian Journal of Zoology 57:2358–2368.
Little, E. L. 1980. The Audubon Society field guide to North American trees. Chanticleer Press, New York, New York, USA.
MacArthur, R. H., and J. W. MacArthur. 1961. On bird species diversity. Ecology 42:594–598.
MacArthur, R. H., and E. O. Wilson. 1967. The theory of island biogeography. Princeton University Press, Princeton, New Jersey, USA.
Marzluff, J. M., F. R. Gehlbach, and D. A. Manuwal. 1998. Urban environments: influences on avifauna and challenges for the avian conservationist. Pages 283–296 in J. M. Marzluff and R. Sallabanks, editors. Avian conservation: research and management. Island Press, Washington, D.C., USA.
McDonnell, M. J., and S. T. A. Pickett. 1990. Ecosystem structure and function along urban–rural gradients: an unexploited opportunity for ecology. Ecology 71:1232–1237.
Meidinger, D., and J. Pojar. 1991. Ecosystems of British Columbia. British Columbia Ministry of Forests, Victoria, British Columbia, Canada.
Melles, S. J. 2001. An examination of urban bird communities in relation to the impact of urbanization and the availability of park space in the lower mainland, BC. Thesis. University of British Columbia, Vancouver, Britissh Columbia, Canada.
Mörtberg, U. M. 2001. Resident bird species in urban forest remnants; landscape and habitat perspectives. Landscape Ecology 16:193–203.
Munyenyembe, F., J. Harris, J. Hone, and H. Nix. 1989. Determinants of bird populations in an urban area. Australian Journal of Ecology 14:549–557.
Odell, E. A., and R. L. Knight. 2001. Songbird and medium-sized communities associated with exurban development in Pitkin County, Colorado. Conservation Biology15:1143–1150.
Ohmart, R. D. 1994. The effects of human-induced changes on the avifauna of western riparian habitats. Studies in Avian Biology 15:273–285.
Pojar, J., and A. MacKinnon. 1994. Plants of coastal British Columbia. Lone Pine Publishing, Vancouver, British Columbia, Canada.
Ralph, C. J., G. R. Geupel, P. Pyle, T. E. Martin, and D. F. DeSante. 1993. Handbook of field methods for monitoring landbirds. USDA Forest Service General Technical Report PSW-GTR-144-www, Pacific Southwest Research Station, Berkley, California, USA.
Reynaud, P. A., and J. Thioulouse. 2000. Identification of birds as biological markers along a neotropical urban–rural gradient (Cayenne, French Guiana), using co-inertia analysis. Journal of Environmental Management 59:121–140.
Rosenberg, K. V., S. B. Terrill, and G. H. Rosenberg. 1987. Value of suburban habitats to desert riparian birds. Wilson Bulletin 99:642–654.
Rottenborn, S. C. 1999. Predicting the impacts of urbanization on riparian bird communities. Biological Conservation 88:289–299.
Saab, V. 1999. Importance of spatial scale to habitat use by breeding birds in riparian forests: a hierarchical analysis. Ecological Applications 9:135–151.
Schaefer, V. 1994. Urban biodiversity. Pages 307–318 in L. E. Harding and E. McCullum, editors. Biodiversity in British Columbia. Environment Canada, Canadian Wildlife Service, Vancouver, British Columbia, Canada.
Sewell, S. R., and C. P. Catterall. 1998. Bushland modification and styles of urban development: their effects on birds in south-east Queensland. Wildlife Research 25:41–63.
Smith, R. J., and J. M. Schaefer. 1992. Avian characteristics of an urban riparian strip corridor. Wilson Bulletin 104:732–738.
Sodhi, N. S., C. Briffett, L. Kong, and B. Yuen. 1999. Bird use of linear areas of a tropical city: implications for park connector design and management. Landscape and Urban Planning 45:123–130.
Soulé, M. E., D. T. Bolger, and A. C. Alberts. 1988. Reconstructed dynamics of urban habitat islands. Conservation Biology 2:75–91.
SPSS. 1996. User’s guide. Base 7.0 for Windows. SPSS, Chicago, Illinois, USA.
Sutherland, G. D., A. S. Harestad, K. Price, and K. P. Lertzman. 2000. Scaling of natal dispersal distances in terrestrial birds and mammals. Conservation Ecology 4(1):16. [Online, URL: http://www.consecol.org/vol4/iss1/art16
Tabachnick, B., and L. S. Fidell. 1996. Using multivariate statistics. Third edition. HarperCollins College Publishers, New York, New York, USA.
ter Braak, C. J. F. 1986. Canonical correspondence analysis: a new eigenvector technique for multivariate direct gradient analysis. Ecology 67:1167–1179.
ter Braak, C. J. F., and P. Šmilauer. 1998. CANOCO reference manual and user’s guide to CANOCO for Windows: software for canonical community ordination. Version 4. Microcomputer Power, Ithaca, New York, USA.
Tilghman, N. G. 1987. Characteristics of urban woodlands affecting breeding bird diversity and abundance. Landscape and Urban Planning 14:481–495.
TRIM (Terrain Resource Information Management). 11993. 1:20000 DEM maps. Ministry of Lands and Parks Surveys and Resources Mapping Branch, Victoria, British Columbia, Canada.
Trzcinski, M. K., F. Lenore, and G. Merriam. 1999. Independent effects of forest cover and fragmentation on the distribution of forest breeding birds. Ecological Applications 9:586–593.
Vale, T. R., and G. R. Vale. 1976. Suburban bird populations in west-central California. Journal of Biogeography 3:157–165.
Vander Haegen, W. M., F. C. Dobler, and D. J. Pierce. 2000. Shrubsteppe bird response to habitat and landscape variables in eastern Washington, U.S.A. Conservation Biology 14:1145–1160.
Vandermeer, J. 1997. The agroecosystem: a need for the conservation biologist’s lens. Conservation Biology 11:591–592.
Weber, W. C. 1972. Birds in cities: a study of populations, foraging ecology and nest-sites of urban birds. Thesis. University of British Columbia, Vancouver, British Columbia, Canada.
World Resources Institute. 1996. The urban environment. World resources Institute, Washington, D.C., USA.
Address of Correspondent:
Stephanie Melles
Zoology Department
University of Toronto
25 Harbord Street
Toronto M5S 3G5 Canada
Phone: (416) 978-7886
stephajm@zoo.utoronto.ca
![]()