|
|
|
Copyright © 2004 by the author(s). Published here under license by The Resilience Alliance.
Go to the pdf version of this article.
The following is the established format for referencing this article:
Hinke, J. T., I. C. Kaplan, K. Aydin, G. M. Watters, R. J. Olson, and J. F. Kitchell. 2004. Visualizing the food-web effects of fishing for tunas in the Pacific Ocean. Ecology and Society 9(1): 10. [online] URL: http://www.ecologyandsociety.org/vol9/iss1/art10/
A version of this article in which text, figures, tables, and appendices are separate files may be found by following this link.
Report Visualizing the Food-Web Effects of Fishing for Tunas in the Pacific Ocean Jefferson T. Hinke1, Isaac C. Kaplan2, Kerim Aydin3, George M. Watters4, Robert J. Olson5, and James F. Kitchell2
1Pacific Fisheries Environmental Laboratory and Joint Institute for Marine and Atmospheric Research; 2Center for Limnology, University of Wisconsin; 3Alaska Fisheries Science Center; 4Pacific Fisheries Environmental Laboratory; 5Inter-American Tropical Tuna Commission
- Abstract
- Introduction
- Methods
- Results
- Discussion
- Conclusion
- Responses to this Article
- Acknowledgments
- Literature Cited
- Appendix 2
We use food-web models to develop visualizations to compare and evaluate the interactions of tuna fisheries with their supporting food webs in the eastern tropical Pacific (ETP) and the central north Pacific (CNP) Oceans. In the ETP and CNP models, individual fisheries use slightly different food webs that are defined by the assemblage of targeted tuna species. Distinct energy pathways are required to support different tuna species and, consequently, the specific fisheries that target different tuna assemblages. These simulations suggest that catches of tunas, sharks, and billfishes have lowered the biomass of the upper trophic levels in both systems, whereas increases in intermediate and lower trophic level animals have accompanied the decline of top predators. Trade-offs between fishing and predation mortality rates that occur when multiple fisheries interact with their respective food webs may lead to smaller changes in biomass than if only the effect of a single fishery is considered. Historical simulations and hypothetical management scenarios further demonstrate that the effects of longline and purse seine fisheries have been strongest in upper trophic levels, but that lower trophic levels may respond more strongly to purse-seine fisheries. The apex predator guild has responded most strongly to longlining. Simulations of alternative management strategies that attempt to rebuild shark and billfish populations in each ecosystem reveal that (1) changes in longlining more effectively recover top predator populations than do changes in purse seining and (2) restrictions on both shallow-set longline gear and shark finning may do more to recover top predators than do simple reductions in fishing effort.
KEY WORDS: ecosystem modeling, food webs, longline fishing, purse-seine fishing, tunas, trophic levels, Pacific Ocean.
Published: March 29, 2004
Evidence for the top-down effects of fisheries on marine ecosystems continues to accumulate (Pauly et al. 1998, Hall 1999, Jackson et al. 2001). Fisheries have been likened to keystone predators (Kitchell et al. 1999) in that the sustained pressure of harvesting can reduce the ecological impacts of the target predators on their prey (Essington et al. 2002). The effects of fishing in marine ecosystems may range from reductions of apex predators (Stevens et al. 2000, Tegner and Dayton 2000, Schindler et al. 2002, Baum et al. 2003) and other economically important fishes (Fogarty and Murawski 1998, Pauly et al. 1998) to the proliferation of unfished or economically unimportant species (Carscadden 2001, Walters and Kitchell 2001, Daskalov 2002) and potential shifts in ecosystem states (Jackson et al. 2001). Recognition of such effects has reinforced recent calls for ecosystem-based management of the world's ocean fisheries (National Marine Fisheries Service 1999, National Research Council 1999).
As evidence for the ecosystem effects of fishing grows, discussions continue as to how the perspectives gained through ecosystem-based research can be incorporated into management strategies (Yaffe 1999, Link 2002). Large system size, limited data, and uncertain knowledge of how fisheries affect entire food webs have hindered the development of useful ecosystem-based management strategies. Consequently, modeling efforts that attempt to link fisheries and food webs have become essential tools for studying ecosystem dynamics and generating hypotheses about the ecosystem effects of fishing (Walters 1986, National Research Council 1999). We use two previously published food-web models (Cox et al. 2002b, Olson and Watters 2003) from two Pacific Ocean ecosystems to compare the effects of the relatively recent expansions of tuna fisheries in the pelagic ocean.
Our primary objective is to compare the potential effects of fishing in two Pacific Ocean ecosystems that have similar food-web structures and fisheries but different histories of fishery development, targets, and bycatch. Appendix 1 contains a brief description of these fisheries and their historical development. We first compare the historical trends in biomass of representative groups that arise from fishery-food web interactions, and we examine the effects that derive from single vs. multiple fisheries. Second, we explore future projections based on hypothetical management objectives that seek to recover shark and billfish populations. The future scenarios we consider arise in response to recent reports of worldwide reductions in the biomass of large pelagic predators (Christensen et al. 2003, Myers and Worm 2003).
An auxiliary goal of this analysis is to better visualize the connections of fisheries to food webs and the effects of fisheries on their food webs. The visualizations used here are intended to highlight the primary connections, defined below, of fisheries to their respective food webs. We also use the visualizations to demonstrate the effects of fisheries, in the context of a food web, that arise from the catches of tunas, sharks, and billfishes.
We used ecosystem models developed for the eastern tropical Pacific (20ºS to 20ºN and 150ºW to the continental shelf of the Americas) and the central north Pacific (0ºN to 40ºN and 130ºE to 150ºW) to explore and compare the connections between the different fisheries and their respective food webs. The spatial extent of the models is displayed in Fig. 1. Each model represents an independent hypothesis about the structure and the potential dynamics of each ecosystem. The groupings, parameterizations, fits to historical time series, and assessments of sources of uncertainty for each model are fully described for the eastern tropical Pacific (ETP) by Olson and Watters (2003) and for the central north Pacific (CNP) by Cox et al. (2002b). We refer the reader to those publications for full details on the models used here.
|
Fig. 1. Map of the Pacific Ocean with the boundaries of the central north Pacific and eastern tropical Pacific systems.
|
Both models were built using the Ecopath with Ecosim (EwE) modeling approach (Christensen and Pauly 1992, Walters et al. 2000) and are representative of average diets and fishery catches from 1993 to 1997 for the ETP and from 1990 to 1998 for the CNP. Together, they summarize a large body of information on the diets, bioenergetics, and life history that is available for pelagic animals in the Pacific Ocean.
The EwE approach facilitates static and dynamic visualizations of ecosystem structure and the ways in which fishing affects this structure. The static Ecopath component is rooted in the thermodynamic principles of mass balance in which production is balanced by losses due to fishing and predation mortality. The mass balance provides a snapshot estimate of the trophic flows that are required to support all the groups and fisheries in the ecosystem. The dynamic Ecosim component describes changes in trophic flows and biomasses caused by changes in fishing effort that occur over the course of fishery development (Fig. 2). We use these alternate representations of the food web to compare static visualizations of food-web configurations from Ecopath output and dynamic visualizations of biomass trends from the Ecosim output.
|
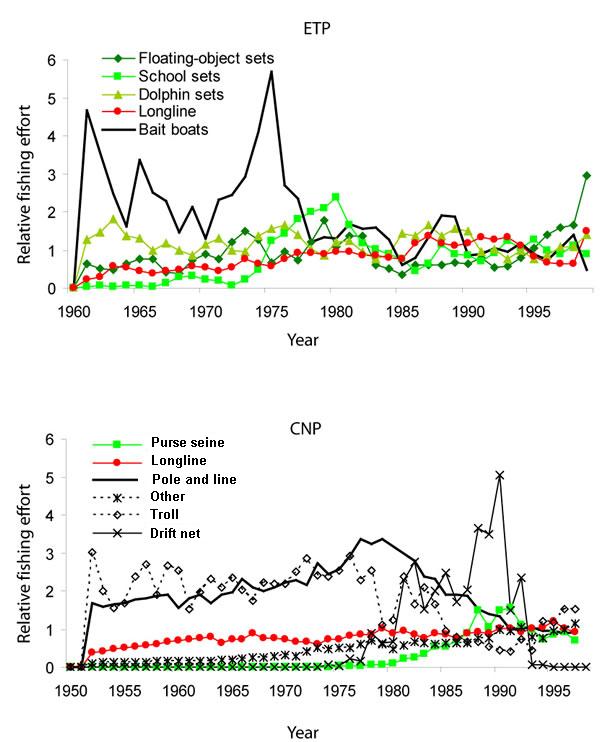
Fig. 2. Indices of historical fishing effort for the eastern tropical Pacific (ETP) and the central north Pacific (CNP) ecosystems. Fishing effort is scaled to the amount of effort expended during the early 1990s. Note that the scaling is applied independently to each fishery, and that it is not appropriate to compare the scaling of effort between fisheries.
|
Static visualizations
We used Ecopath estimates of the mass-balanced food webs to trace the primary trophic linkages from the major fisheries to the primary producers at the base of each food web. The major fisheries were defined as those that cumulatively accounted for the top 95% of the 1990s catch in the model. The primary targets were defined as the set of species or groups in the model that accounted for the top 95% of each major fishery's catch. In turn, all remaining linkages in each food web were identified by the set of prey that accounted for 95% of the diet of the targeted predators. This method was applied for each species or group included, down to the primary producers, to produce fishery-specific food webs that we termed "fishery food webs." Our arbitrary cutoff of 95% highlights the primary energy flows within the model that support those fisheries with the highest yields. The ELVIZ software used to produce the visualizations and comparisons (Aydin et al. 2002) is available from the Alaska Fisheries Science Center of the National Marine Fisheries Service.
Sharks and billfishes were not well represented in the fishery food webs (see Results), because these fishes were not large components of catches in the 1990s. The bycatch of these apex predators is a special concern for fisheries management agencies in the Pacific Ocean (Bailey et al. 1996, Inter-American Tropical Tuna Commission 2002a), owing, in part, to life-history attributes that render their populations susceptible to overfishing (Stevens et al. 2000, Schindler et al. 2002). To describe the ETP and CNP food webs of important bycatch species, which we termed "bycatch food webs," we separately applied the same 95% approach to the billfish and shark groups whose primary mortality sources included fisheries.
We computed a measure of connectance to provide a quantitative comparison of each fishery and bycatch food web with the respective full food web for the ETP and CNP. We define connectance here as the ratio of the number of links in each fishery food web or bycatch food web to the total number of links in a full (95% criteria) food web that includes all fisheries. For example, if a fishery food web contains only 10 links and the full food web contains 20, the connectance for the fishery food web is 0.5.
Dynamic visualizations and simulationsWe used the Ecosim results from two sets of fishing scenarios for both the ETP and CNP to develop dynamic visualizations of fishery-induced variations in the food web. First, the model was forced with time series of historical fishing effort (Fig. 2) that allowed us to compare the effects of historical fishing patterns on food-web components. We animated the effect of the combined fishing effort from all fisheries to produce a food-web movie of the potential system-wide effects of historical fishing in each area. The second set of scenarios compares the combined effects of multiple gears to the effects of individual fisheries. We isolated the effects of purse-seine and longline fisheries in both the ETP and CNP by comparing simulations with all fishery effort to simulations without the fishing effort from either purse seines or longlines. The difference between the simulations was considered to represent the effect of the excluded fishery, given the existence of all other fisheries. This method allowed us to investigate more closely the food-web effects of an individual fishery that may be masked when all fisheries are considered in aggregate.
Alternative management scenariosConservation of marine apex predators such as sharks and billfish is a pressing global problem (Christensen et al. 2003, Myers and Worm 2003). Although the full extent of the depletion of these predators in the Pacific Ocean is a subject of debate (Hampton et al. 2003, Walters 2003), it is reasonable to expect that future management actions will seek to increase the abundance of these species. Management options for rebuilding predator populations could include changing fishing gear, restricting the retention of certain bycatch, or reducing the fishing effort of certain fleets. Here we investigated five examples of these management policies. For each example, we started with the base 1990s models and projected forward for 30 yr under the new management scenario. The catches and mortality rates for all species and the fishing effort of all fisheries not affected by the management action were maintained at their average 1990s levels. Using food-web diagrams, we illustrate the direct effects of lowered fishing mortality and the indirect effects related to changes in predation and competition after the 30-yr implementation of the new fishing policy.
One management option for increasing billfish stocks would be to require longline fishermen to set all hooks deeper than 120 m. Based on catch-at-depth information, an effective ban on shallow-set hooks could reduce the catch of marlins (Hanamoto 1974, Boggs 1992, Nakano et al. 1997). The results from Boggs (1992) suggest that eliminating the shallow hooks from longlines would reduce longline fishing mortality for marlin, other billfishes except swordfish, yellowfin, and bigeye tuna by 54, 42, 8, and 2%, respectively. We applied these reduced longline fishing mortality rates to billfish and tuna mortality in both the ETP and CNP. The reduced mortality scenario is similar to the ecological economics analysis by Kitchell et al. (2004) that focuses specifically on responses to longline modification in the CNP.
A second management option might be to ban the finning of sharks. Currently, only the fins of most pelagic sharks are retained (McCoy and Ishihara 1999, Ito and Machado 2000). Sharks that do not have their fins removed are often alive (Hoey 1996, Lee and Brown 1998) and likely to survive after being released. The United States has already banned shark finning (National Archives and Records Administration 2000), but the other major fishing nations have not. We tested the impact of a Pacific-wide ban on shark finning from longline vessels. We projected the base ecosystem model forward with a 73% reduction in longline fishing mortality on all sharks (Hoey 1996). We also evaluated the effects of a management strategy that combines the shark-finning ban and the elimination of shallow-set longline hooks.
The crudest management option is simply to cut longline or purse-seine fishing effort. Reducing longlining effort diminishes the bycatch of marlins and sharks, and reducing purse-seine fishing may increase the abundance of prey available to apex predators. Therefore, we ran two simulations, one with a 50% reduction in longline effort and the other with a 50% reduction in purse-seine effort.
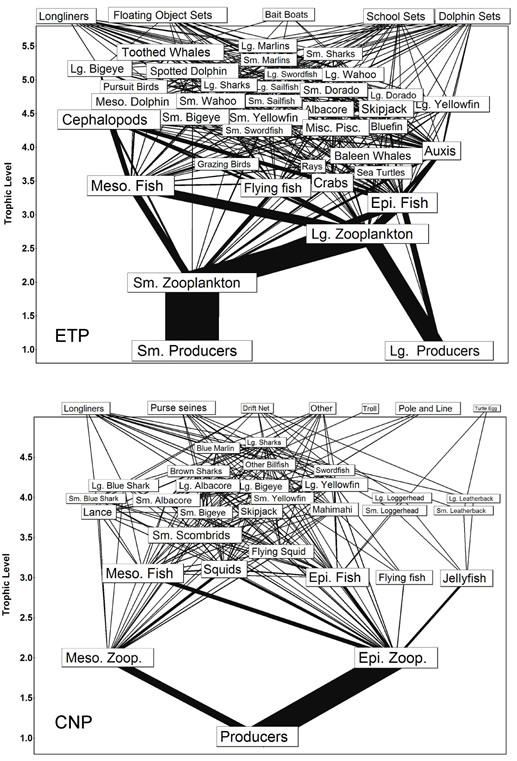
The full food webs of the eastern tropical Pacific (ETP) and central north Pacific (CNP) models are presented in Fig. 3. Each model incorporated 31 or more biomass groups and five or more fisheries. The trophic structure of both systems above primary producers was modeled in the same way. The highest resolution occurred in the upper trophic levels, for which data were most abundant. The lower trophic levels were generally more aggregated. In general, the display in Fig. 3 was not amenable to a simple interpretation of interactions between fisheries and their food webs. We were able to simplify our portrayal of these complex food webs by focusing only on single fisheries and the food webs that support them.
|
Fig. 3. Full food webs for the eastern tropical Pacific (ETP) and central north Pacific (CNP) models. These were developed from the models described by Olson and Watters (2003) and Cox et al. (2002b), respectively. Fisheries are not positioned according to their estimated trophic levels.
|
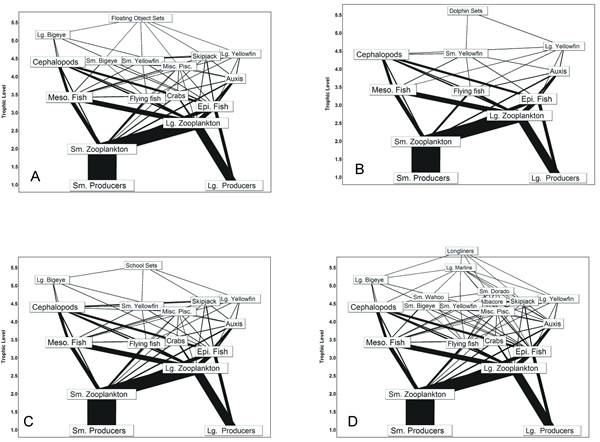
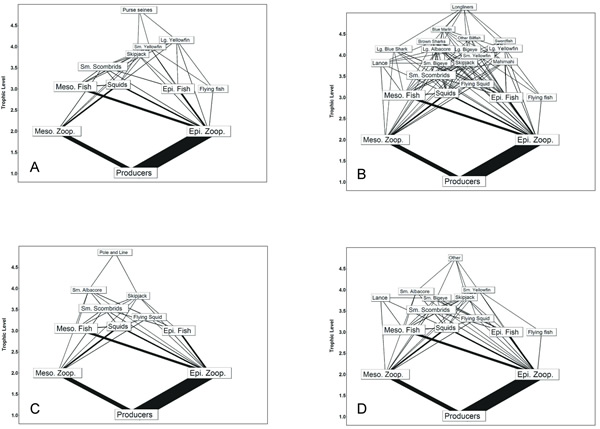
In the ETP model, the purse-seine and longline fisheries together accounted for more than 95% of the total catch during the 1990s. Of the three purse-seine fisheries, the floating-object fishery accounted for 28% of the total catch, and the primary harvest included bigeye, yellowfin, and skipjack tunas. The floating-object fishery depended on a relatively complex food web (Fig. 4, Table 1). In contrast, the dolphin-set fishery, the second-largest fishery in terms of catch, caught primarily large and small yellowfin and was supported by the smallest fishery food web. The school-set fishery food web was intermediate to the other purse-seine food webs (Table 1). The longline fishery food web was larger than any of the purse-seine webs (Fig. 4). The diversity was caused by a small catch of marlins, which resulted in the highest connectance for all ETP fishery food webs (Table 1). For all fisheries, the target species, i.e., yellowfin, skipjack, and bigeye tunas, fed primarily on squids, flying fishes, small scombrids such as Auxis spp., and epipelagic and mesopelagic fishes.
|
Fig. 4. Fishery food webs supporting the fisheries that accounted for at least 95% of total catch in the eastern tropical Pacific (ETP) during the 1990s. Fishing gear included floating-object sets (A), dolphin sets (B), school sets (C), and longline (D). Fisheries are positioned according to their estimated trophic levels.
|
|
Table 1. Comparison of the four major fishery and the bycatch food webs for the eastern tropical Pacific (ETP) and central north Pacific (CNP). For the fishery food webs, the number of groups caught is a count of the groups that account for 95% of the fishery catch in the 1990s. For the bycatch food webs, the number of groups caught is a count of the shark or billfish groups whose primary mortality (95% of total) includes bycatch. The number of groups caught defines the top of each food web. The fraction of total catch is calculated as the fraction of the total system catch that is attributed to the catch or bycatch of the number of groups caught. Connectance is calculated as the fraction of links from the full food webs that were present in the fishery or bycatch food webs.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
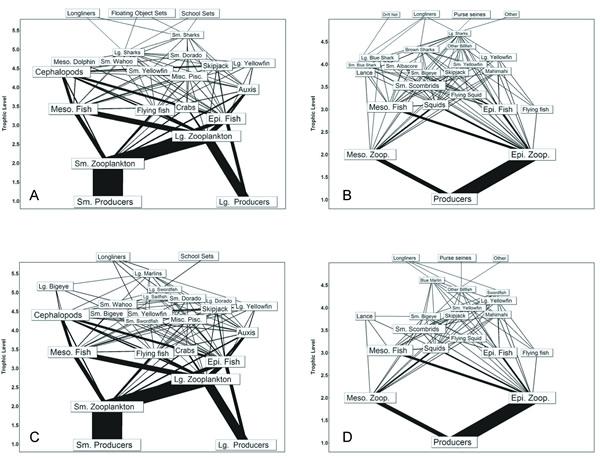
In the CNP model, the purse-seine, longline, pole-and-line, and "other" fisheries accounted for more than 95% of the total catch during the 1990s (Fig. 5). The largest fishery in terms of total catch (50%) was the purse-seine fishery. Note that, for the CNP model, no distinction was made between alternative methods of purse-seine deployment. The CNP purse-seine food web was relatively simple compared to the ETP purse-seine food webs, with the major catches comprising skipjack and yellowfin tunas (Fig. 5). As in the ETP model, the longline food web in the CNP was the most complex, and a small catch of both sharks and billfishes with broad diets contributed strongly to both higher diversity and higher connectance (Table 1). The "other" fishery took 10% of the total catch, but the catch of four tuna groups contributed to the second-highest connectance for the fishery food webs in the CNP. The pole-and-line fishery food web was the smallest of the CNP fishery food webs. The primary target species, i.e., skipjack, yellowfin, and bigeye tuna, of the main fisheries in the CNP also fed primarily on small scombrids, squids, flying fishes, and small epipelagic and mesopelagic fishes. Thus, the lower trophic levels that support the tuna fisheries in the CNP were analogous to those of the ETP.
|
Fig. 5. Fishery food webs supporting the fisheries that accounted for at least 95% of total catch in the central north Pacific (CNP) during the 1990s. Fishing gear included purse seine (A), longline (B), pole and line (C), and other (D). Fisheries are positioned according to their estimated trophic level.
|
For both models, the differences in fishery food-web configuration were evident primarily above trophic level 3 (Fig. 5). However, despite general differences in the primary assemblage of catch for each fishery, there was some overlap of catch species across all the fisheries within each system, and the primary pathways of energy flow through highly aggregated trophic levels 1–3 were identical in both the ETP and CNP models. For instance, the ETP purse-seine food webs could be considered nested versions of the floating-object set food web, and large yellowfin were caught by all the major fisheries in the ETP (Fig. 4). The subtle differences in the composition of catch among fisheries, however, emphasized that different energy pathways were required to link different fisheries to the lower trophic levels.
Shark and billfish bycatch food webs were relatively complex (Fig. 6, Table 1). Compared with the fishery food webs, the bycatch food webs were defined by fewer groups caught and represented a smaller proportion of total catch, yet still had a connectance comparable to or higher than those of longline food webs (Table 1). Indirect interactions between fisheries and their principal bycatch species were also illustrated in the bycatch food webs. For example, longline and purse-seine fisheries caught marlin and the primary prey of marlin, i.e., yellowfin, bigeye, and skipjack (Figs. 4 and 5), removing marlin biomass directly and harvesting the available prey base for remaining marlin. Such bycatch and fishery food webs illustrate the potential for fisheries to influence individual groups through multiple pathways, not simply via direct removal by the fisheries.
|
Fig. 6. Shark and billfish bycatch food webs for the eastern tropical Pacific (A and C) and the central north Pacific (B and D). Shark bycatch webs are in the upper panels (A and B), and billfish bycatch webs are in the lower panels (C and D). Fisheries are not positioned according to their trophic levels.
|
Dynamic visualizations and simulations
Once fishing began in each pelagic ecosystem, immediate declines in the biomass of tunas, sharks, and billfishes in both the ETP (Fig. 7) and CNP (Fig. 8) were observed in both models. These reductions in predator biomass were apparently accompanied by reciprocal increases in the biomass of lower trophic levels. For example, in the ETP model in Fig. 7, the declines in upper trophic levels were mirrored by increases in nontarget Auxis and dorado (Coryphaena spp.). Further, the CNP model predicted analogous increases in the small scombrid and mahi-mahi groups that were mirrored by declines in flying fishes and squids (Fig. 8). The aggregate effect of tuna fishing in both models appears to have been a shift in the distribution of biomass from upper-level predators to their prey.
|
Fig. 7. Animation of the effects of historical fishing effort in the eastern tropical Pacific (ETP). The animation begins a year before the start of large-scale industrial fishing. Each animation frame represents 1 yr and depicts the Ecosim estimate of the percent change in biomass from the estimated prefishing biomass for all groups. Green shading indicates increases in biomass, and red shading indicates decreases in biomass. Our estimate of prefishing biomass was obtained from model projections under no fishing effort (F = 0) conditions. We assume that the equilibrium biomasses obtained under the F = 0 simulations approximate the biomasses in an unfished system. The relative fishing effort, i.e., relative to the fishing effort expended in the 1990s, of each tuna fishery is annually updated in the animation frames. Most of the variation in the fishing effort of the largest fleets varies between 0 and 1.5, so we capped the y-axis to highlight that variability. The full range of effort values is presented in Fig. 2.
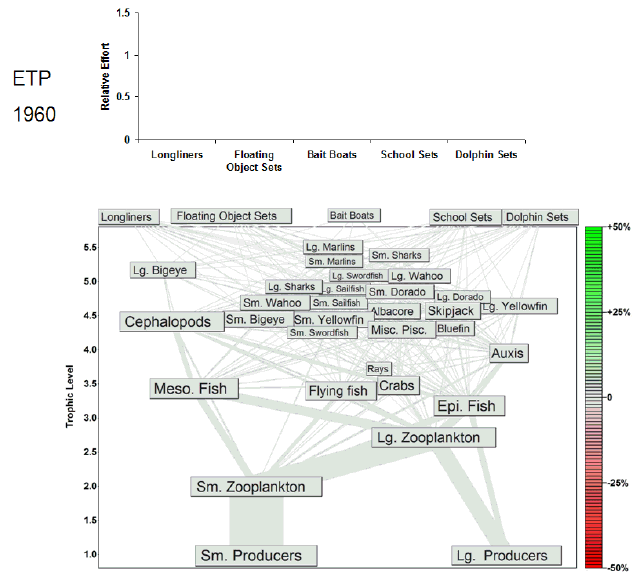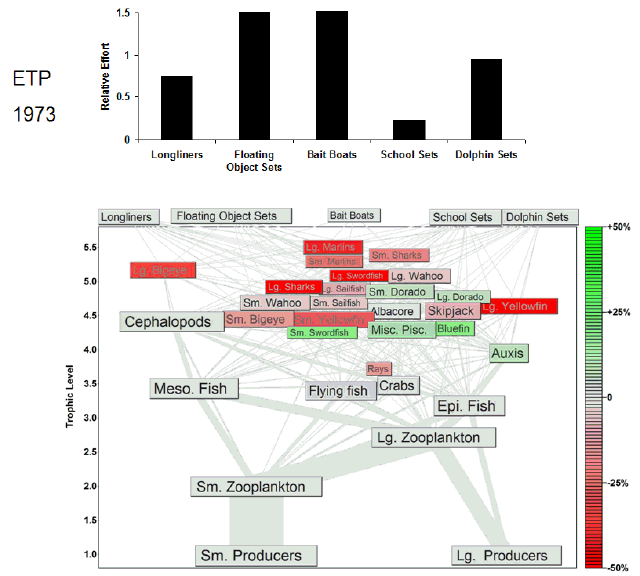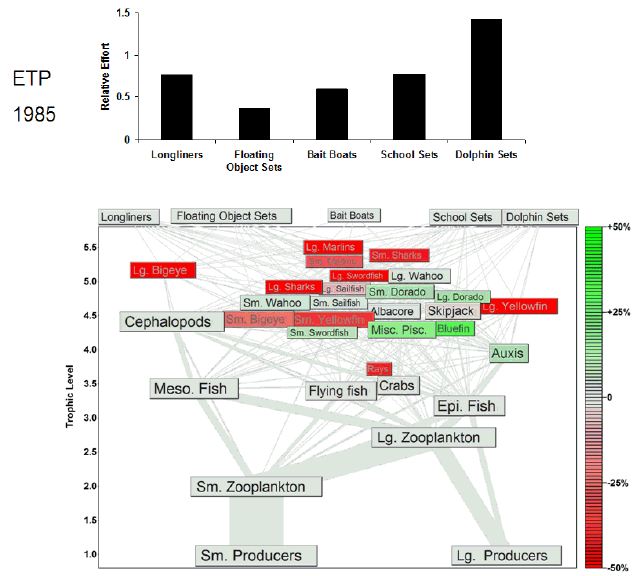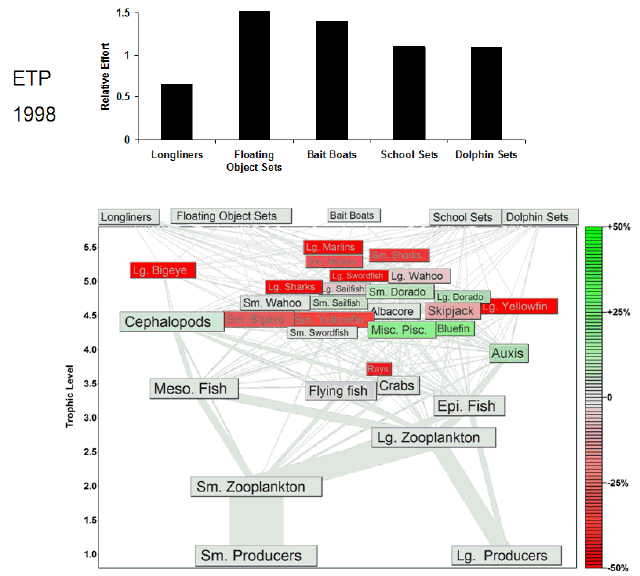
|
|
Fig. 8. Animation of the effects of historical fishing effort in the central north Pacific (CNP). The animation begins a year before the start of large-scale industrial fishing. Each animation frame represents 1 yr and depicts the Ecosim estimate of the percent change in biomass from the estimated prefishing biomass for all groups. Green shading indicates increases in biomass, and red shading indicates decreases in biomass. Our estimate of prefishing biomass was obtained from model projections under no fishing effort (F = 0) conditions. We assume that the equilibrium biomasses obtained under the F = 0 simulations approximate the biomasses in an unfished system. The relative fishing effort, i.e., relative to the fishing effort expended in the 1990s, of each tuna fishery is annually updated in the animation frames. Most of the variation in the fishing effort of the largest fleets varies between 0 and 1.5, so we capped the y-axis to highlight that variability. The full range of effort values is presented in Fig. 2.

|
When all fisheries operated simultaneously, the effects of single fisheries were not readily detectable in Figs. 7 and 8. The large, early declines in sharks and billfishes in the CNP occurred in the absence of purse seining and appeared to result primarily from longline fishing (Fig. 8). However, bycatch from the "other" fishery also contributed to the catch of these apex predators (Fig. 6). When we isolated the effects of purse seining and longlining on the food webs of each model, the direct and indirect effects attributable to individual fisheries emerged.
As stated previously, the effect of an individual fishery was inferred by comparing simulations with all fisheries to simulations with all fisheries minus the particular fishery of interest. Purse seining and longlining, respectively, had system-wide effects that were similar between the ETP and CNP models. Purse-seine fisheries in the ETP reduced the biomass of bigeye, yellowfin, and sharks (Fig. 9), whereas CNP purse seining reduced skipjack biomass by 50% (Fig. 10). The biomass of the small scombrids group that were functionally similar to the Auxis spp. in the ETP indirectly increased in both systems (Figs. 9 and 10). The increase in small scombrids was presumably the result of decreased predation by the targeted tunas, which are important predators of small scombrids. The effects of the model longline fisheries were nearly identical in both food webs. Reductions in the biomass of bigeye, albacore, marlins, and sharks occurred, and the negative trends persisted throughout (Figs. 9 and 10). The model results suggested that the effects of longlining were mainly direct and strongest at the upper trophic levels, whereas the primary indirect response of the lower trophic levels was apparently to purse-seine effort.
|
Fig. 9. Biomass predictions in the eastern tropical Pacific (ETP) from the effect of all fisheries, longliners only, and purse seiners only. Trends are arrayed roughly according to trophic levels with a typical purse-seine food chain on the right and a longline food web on the left. The groups included were based on fishery and bycatch food webs and the availability of analogous groups in the central north Pacific.
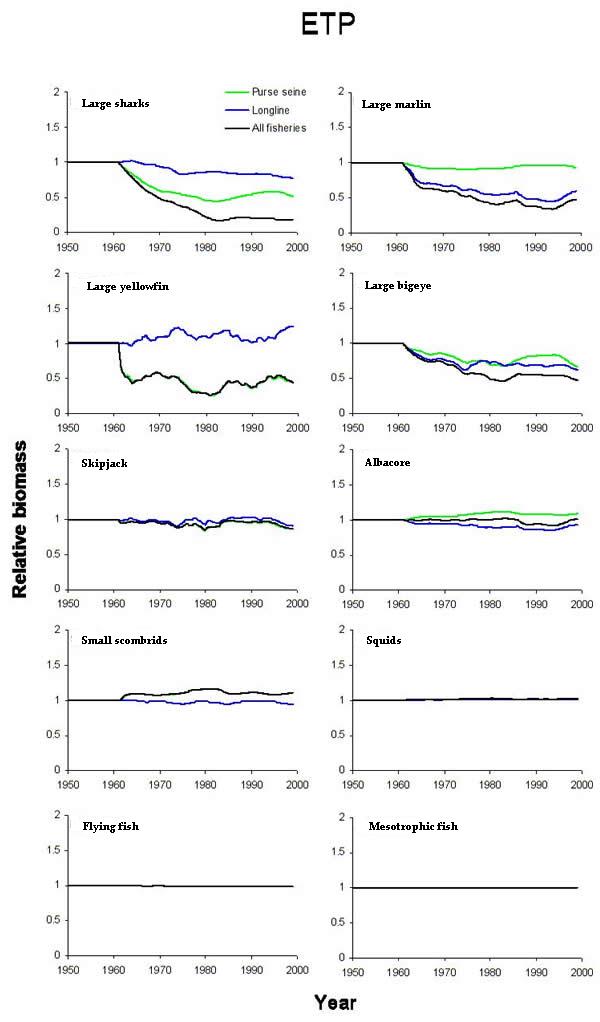
|
|
Fig. 10. Biomass predictions in the central north Pacific (CNP) from the effect of all fisheries, longliners only, and purse seiners only. Trends are arrayed roughly according to trophic level. The groups are analogous to those in Fig. 9.
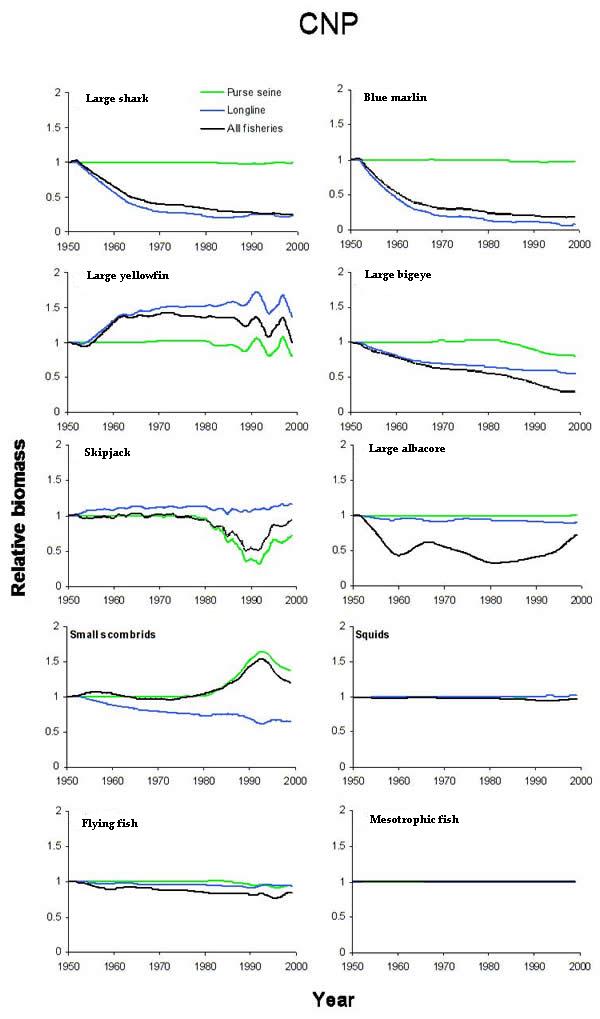
|
The predicted effects of the single fisheries often led to higher levels of biomass reduction than the combined effect of multiple, concurrently operating fisheries. For example, CNP skipjack responded strongly to the purse-seine fishery. If purse seining had been the only fishery in the CNP, skipjack biomass might have been reduced below the level that resulted from both longlining and purse seining. Similar situations were observed for small scombrids, blue marlin, and large sharks in the CNP and for albacore in the ETP (Figs. 9 and 10). In some cases, the aggregate fishery effect was often closely approximated by the effect of only one fishery. The declines in marlin and bigeye were closely approximated by longline fisheries alone in both models. Skipjack and small scombrids biomass trends were driven mainly by purse seining (Figs. 9 and 10).
Alternative management scenariosIn the ETP and CNP, the banning of shallow longline gear and shark finning led to the recovery of the marlin and shark populations, respectively. In the ETP, the ban on shallow longline gear led to increases in marlin biomass of more than 60% (Table 2, Fig. 11A). In the CNP, the ban on shallow longline gear allowed blue marlin biomass to nearly double, and "other billfish," mostly striped marlin, increased by more than 50% (Figure 12A). Increased marlin stocks then caused minor reductions in other food-web components, notably by preying on and competing with yellowfin tuna and competing with sharks in the ETP. The rest of the food web did not respond strongly to the alteration of the longline fishing strategy.
|
Table 2. Percent change in biomass of select groups owing to the hypothetical changes in fishing strategies or fishing effort.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
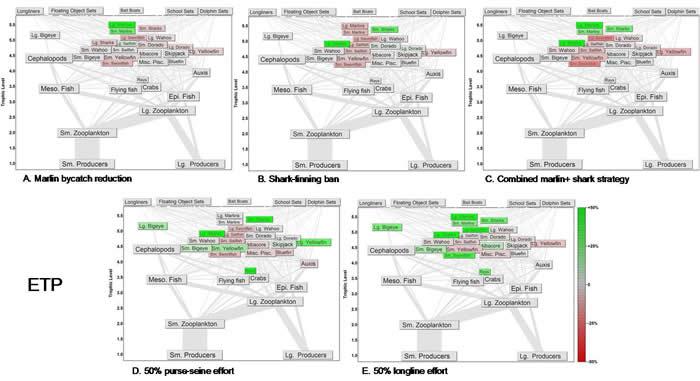
Fig. 11. Response of the food web in the eastern tropical Pacific (ETP) after 30 yr of alternative management strategies for apex predator recovery.
|
|
Fig. 12. Response of the food web in the central north Pacific (CNP) after 30 yr of alternative management strategies for apex predator recovery.

|
The ban on shark finning caused increases in all shark groups. Shark biomass doubled in the ETP within 30 yr (Fig. 11B). In the CNP, adult blue sharks, large sharks, and brown sharks all increased by more than 60% (Table 2, Fig. 12B). An indirect effect of the finning ban in both systems was that shark predation increased on marlins and mid-trophic-level species such as tunas. Marlin in both models declined by roughly 10% under the shark finning ban. The rest of the food web did not respond strongly to the change in shark fishing mortality.
Combining the finning ban and the ban on shallow longline gear led to a general recovery of marlin and sharks. In this scenario, marlin recovered to levels slightly lower than if only shallow gear had been banned. The ETP shark responses to the combined ban were generally lower than if only finning had been banned (Fig. 11C). Shark recovery in the CNP was similar to shark recovery under the finning ban only (Fig. 12C). Yellowfin biomass in both systems declined more strongly under the combined scenario than for either alternative fishing strategy alone; this was caused by increased predation by both marlin and sharks on yellowfin. The rest of the food web did not respond strongly to the combined finning ban and shallow-gear ban.
Reducing purse-seining effort by 50% had no positive effects on marlin in either food web. Sharks increased about 80% in the ETP, where they are a large component of bycatch in purse-seine sets (Fig. 11D). CNP sharks were not influenced by the purse-seine reduction (Fig. 12D). In both systems, tunas responded most strongly to the reduction in purse-seine effort (Table 2). In turn, this led to declines of about 20% in the small scombrids of the CNP because of higher predation by tunas (Fig. 12D).
Reducing longlining effort by 50% caused increases in both billfish and shark biomasses in both food webs (Figs. 11E and 12E). In most cases, however, the group-specific increases were not as strong as those resulting solely from the shark-finning ban, the shallow-gear ban, or the combined strategy. Under the reduced longline effort scenario, bigeye biomass increased because of lower fishing mortality, whereas yellowfin biomass decreased slightly because of higher predation rates.
In the eastern tropical Pacific (ETP) and the central north Pacific (CNP), the development of pelagic tuna fisheries has caused similar changes in the overall structure of the food webs. Simulations suggest that the catches of tunas, sharks, and billfishes have lowered the biomass of the upper trophic levels in both systems, whereas increases in animal numbers at the intermediate and lower trophic levels have accompanied the declines of top predators. Historical simulations and hypothetical management scenarios further demonstrated that the effects of longline and purse-seine fisheries have been strongest in the upper trophic levels, but that lower trophic levels may respond more strongly to purse-seine fisheries. The apex predator guild responded most strongly to longlining. The differences in the food-web effects resulting from longline and purse-seine fisheries further identify trade-offs that could help guide ecosystem-based management actions.
The pathways connecting ETP and CNP fisheries to their primary food webs depend on the different assemblages of tunas that are caught by each fishery, thereby creating fishery-specific food webs. The overlap of primary forage groups among the fishery food webs create conditions that allow the effect of multiple fisheries, each acting on different assemblages of tunas and other large predators, to concentrate indirect effects on the same set of forage groups. A comparison of the ecosystem effects of the historical development of fisheries showed that middle and lower trophic levels in the ETP and CNP may be sensitive to reductions in upper-level predator biomass. In both systems, it was shown that biomass reductions in the upper trophic level were accompanied by increases in the biomass of the lower trophic levels, a finding that is similar to the growing evidence from other fished ecosystems (e.g., Lilly et al. 2000, Carscadden et al. 200l).
The sensitivity of the lower trophic levels in our models was often minor, and these levels frequently exhibited a change of less than 20% in their biomass. As discussed below, given the host of uncertainties associated with ecosystem models, responses of such magnitude should be viewed with some skepticism. However, our results do suggest that the potential for indirect interactions exists and that the primary food webs for each fishery can be relatively simple. Therefore, the propagation of fishery effects throughout the food web should not be dismissed as unlikely because of apparent food-web complexity. Changes in lower trophic levels could happen if the perturbations to the upper trophic levels are sufficiently large, such as those caused by the expansion and intensification of the fishing effort over the last 50 yr.
The simulations also suggested that an increase in biomass caused indirectly by one fishery may buffer the declines driven directly by another fishery. Generally, we expect fishing to increase the total mortality of a species. However, if one fishery tends to remove the predators of the target species of another fishery, then the balance of increased fishing mortality and reduced predation mortality may lead to little net change in biomass for this target species. For example, skipjack in the CNP decline due to targeted fishing by the purse-seine fishery. However, skipjack tend to increase when their predators, i.e., bigeye, are caught by the longline fishery. Combining longline and purse-seine fisheries leads to an intermediate result, which is less extreme than under either individual fishery. For groups that are caught by multiple fisheries, the additive effect of increased fishing mortality likely outweighs any potential declines in predation mortality, and stronger reductions in biomass under combined fisheries than under individual fisheries can result. For example, large bigeye in both systems were targeted by multiple fisheries (Figs. 4 and 5), and their response to combined fishing by all gears was larger than their response to either longlining or purse seining alone.
Alternative management strategies to rebuild shark and billfish populations in each ecosystem revealed differences between (1) purse seines and longlines in terms of their effectiveness for recovering top predators and (2) gear modifications and bycatch laws vs. effort reductions in longlining. Our results suggested that decreases in longline effort made it possible to rebuild the ETP and CNP shark and billfish populations, whereas the reduction in purse-seine effort had little influence on the recovery of top predators in both systems. Thus, the role of longlining generally appears to be more critical to the conservation of sharks and billfish in the Pacific.
However, identifying management options for future longline fisheries to achieve shark and billfish recovery was not a straightforward task. Our simulations suggested that a simple reduction of longline effort could allow a larger number of populations to rebuild, but the overall recovery of sharks and billfishes might be less marked and at potentially higher costs to the fishery than under a more conventional management strategy that emphasizes gear modifications and bycatch retention practices. Although the combined prohibition of shallow gear and shark finning was more effective at recovering marlin and sharks than a reduction of longlining effort, few other species benefited. Under both management strategies, increased predation may initiate further declines in yellowfin biomass. Furthermore, enacting a ban only on shark finning may allow for a recovery of shark populations, but may also cause an unintended decline in marlin populations; this represents a bitter twist to a policy intended to conserve pelagic apex predators. Our simulations also suggested that the largest changes in the biomass of any billfish or shark group in response to any fishing strategy were only slightly greater than a doubling of biomass and, more typically, less than an 80% increase. If recovery goals mandate higher equilibrium biomasses for top predators (e.g., Myers and Worm 2003), then a combination of stronger restrictions on gear deployment, bycatch retention, and fishing effort may be required to allow predator stocks to rebuild.
CaveatsWhen considering an analysis of this type, it is important to take into account the possible shortcomings of a food-web modeling approach. The authors of the original ETP (Olson and Watters 2003) and CNP (Cox et al. 2002b) models both provide thorough discussions of the uncertainties that they felt most affected their initial analyses. Below, we discuss some of those uncertainties with respect to the comparative inferences of this paper.
Ecosystem analyses often rely on an incomplete knowledge of the ecological interactions among a diverse group of species. The models we used represent a hypothesis about system structure and the potential dynamics that arise as a result of that structure. Specifically, the model structure depends on information about diets, catch, and bycatch. Diet data are particularly difficult to quantify in pelagic systems. As diet data and other ecological information for more taxa become better known, the configuration and dynamics of these models will likely change, along with our understanding of how these systems function. In the ETP model, Olson and Watters (2003) found that trophic flows were most sensitive to parameter estimates for Cephalopods and Auxis spp. for which few to no empirical data were available. However, in dynamic simulations under historic patterns of fishing effort, they found that the sensitivity of the model to these two groups affected levels of predicted biomass throughout the food web, but not trends in biomass (Olson and Watters 2003). As such, our comparisons of the magnitude of fishery effects in the ETP and CNP should be viewed with some skepticism, but it remains probable that the overall food-web trends that arise as a result of fisheries effects are reasonable.
Another large source of uncertainty in our comparisons derives from the animal groupings, which range from specific to generic in each model. For some groups, comparisons are fairly straightforward, because a single species was modeled and was common to both models, e.g., yellowfin or bigeye tunas. For other groups such as large sharks, the ETP group did not contain the same species as the CNP group. In general, species with different life histories were often lumped together into functional guilds. The guild members in each model were often reasonably similar and fulfilled functionally equivalent roles, but they were not exact replicas. Thus, it would be false to assume that all the species included in a larger grouping responded in exactly the same manner as the guild, neither within nor between the modeled systems.
Expectations of how food webs will respond to fishing may further depend on the nature of physical forcing. Specifically, the ability to identify the effects of fishing, particularly at lower trophic levels, will almost certainly depend on the strength and frequency of bottom-up forcing that the food web experiences (Watters et al. 2003). Our results from considering only the effects of fisheries suggest that lower trophic levels may respond to fishing, but disentangling environmental signals from fishery and predation signals in data is a task fraught with considerable challenges for management and future studies based on an ecosystem perspective.
A final matter of concern is the vast spatial extent of the modeled ecosystems. The large-scale perspective adopted by approaches that take into account the entire food web does restrict the ability of these models to capture important local interactions among smaller groups of species or between fisheries and their food webs in localized areas. Further, the ETP and CNP models together span the Pacific Ocean (Fig. 1), and there is no true biological boundary that separates the western ETP from the eastern CNP. As such, if animals occupy both regions, it is uncertain which of the two model hypotheses, if not an alternative model, might take precedence. We do not know how important such overlap might be to these ecosystems.
We conclude by summarizing the emergent themes of this comparative analysis of two pelagic ecosystems. First, both pelagic ecosystems were similarly structured, but the individual fisheries that operate in each system rested on dissimilar food webs. Our visualizations identified the primary target and bycatch groups and the important food resources used by those targets. These simplified food webs could be used to distinguish and, perhaps, monitor fishery effects in the future. Second, the main ecosystem effects of tuna fishing in both the ETP and CNP models were similar and can be characterized by (1) the reduction of apex predators, analogous to recent reports of world wide trends (Dayton et al. 2002, Baum et al. 2003), and (2) the increased biomass of some prey species that resulted from the release of predation. The food-web animations presented in Figs. 7 and 8 highlighted these effects. The strongest perturbations to these ecosystems were the simultaneous development and growth of multiple, concurrent tuna fisheries. However, the effects of longlining appear strongest at the top of the food web, whereas the purse-seine fisheries caused larger changes in biomass at lower trophic levels. Attempts to recover top predators by reducing fishing mortality via longline gear modifications and by stricter regulations on shark finning may be as effective as simple reductions in longline fishing effort.
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a comment, follow this link. To read comments already accepted, follow this link.
This work was conducted as part of the Marine Management Working Group supported by the National Center for Ecological Analysis and Synthesis, a center funded by the NSF (Grant #DEB-0072909), the University of California, and the Santa Barbara campus. We thank S. Harley and two anonymous referees for constructive reviews of a previous draft. JTH and JFK were supported by a grant from Wisconsin Sea Grant. JFK and ICK were supported by a grant from the NSF Biological Oceanography Program. The results and interpretations presented in this paper are the views of the authors and are not intended to reflect the opinions and policies of NOAA.
Aydin, K. Y., V. V. Lapko, V. I. Radchenko, and P. A. Livingston. 2002. A comparison of the eastern Bering and western Bering Sea shelf and slope ecosystems through the use of mass-balance food web models. NOAA Technical Memorandum NMFS-AFSC-130. NOAA Fisheries, Silver Spring, Maryland, USA.
Bailey K., P. Williams, and D. G. Itano. 1996. Bycatch and discards in central and western Pacific tuna fisheries: a review of SPC data holdings and literature. Ocean Fisheries Programme Technical Report Number 34. South Pacific Commission, Noumea, New Caledonia.
Baum, J. K., R. A. Myers, D. G. Kehler, B. Work, S. J. Harley, and P. A. Doherty. 2003. Collapse and conservation of shark populations in the Northwest Atlantic. Science 299:89-392.
Boggs, C. H. 1992. Depth, capture time, and hooked longevity of longline-caught pelagic fish: timing bites of fish with chips. Fisheries Bulletin 90:642-658.
Calkins, T. P. 1980. Synopsis of biological data on the bigeye tuna, Thunnus obesus (Lowe, 1839), in the Pacific Ocean. Pages 213-260 in W. H. Bayliff, editor. Synopses of biological data on eight species of scombrids. Inter-American Tropical Tuna Commission, La Jolla, California, USA.
Carscadden, J. E., K. T. Frank, and W. C. Leggett. 2001. Ecosystem changes and the effects of capelin (Mallotus villosus), a major forage species. Canadian Journal of Fisheries and Aquatic Sciences 58:73-85.
Christensen, V., and D. Pauly. 1992. Ecopath II—a software for balancing steady-state models and calculating network characteristics. Ecological Modelling 61:169-185.
Christensen, V., S. Guenette, J. J. Heymans, C. J. Walters, R. Watson, D. Zeller, and D. Pauly. 2003. Hundred-year decline of North Atlatic predatory fishes. Fish and Fisheries 4:1-24.
Cox, S. P., S. J. D. Martell, C. J. Walters, T. E. Essington, J. F. Kitchell, C. Boggs, and I. Kaplan. 2002a. Reconstructing ecosystem dynamics in the central Pacific Ocean, 1952-1998. I. Estimating population biomass and recruitment of tunas and billfishes. Canadian Journal of Fisheries and Aquatic Sciences 59:1724-1735.
Cox, S. P., T. E. Essington, J. F. Kitchell, S. J. D. Martell, C. J. Walters, C. Boggs, and I. Kaplan. 2002b. Reconstructing ecosystem dynamics in the central North Pacific Ocean, 1952-1998. II. A preliminary assessment of the trophic impacts of fishing and effects on tuna dynamics. Canadian Journal of Fisheries and Aquatic Sciences 59:1736-1747.
Daskalov, G. M. 2002. Overfishing drives a trophic cascade in the Black Sea. Marine Ecology Progress Series 225:53-63.
Dayton, P. K., S. Thrush, and F. Coleman. 2002. Ecological effects of fishing in marine ecosystems of the United States. Pew Oceans Commission, Arlington, Virginia, USA.
Edwards, E. F., and P. C. Perkins. 1998. Estimated tuna discard from dolphin, school, and log sets in the eastern tropical Pacific Ocean, 1989-1992. Fishery Bulletin 96:210-222.
Essington, T. E., D. E. Schindler, R. J. Olson, J. F. Kitchell, C. Boggs, and R. Hilborn. 2002. Alternative fisheries and the predation rate of yellowfin tuna in the eastern Pacific Ocean. Ecological Applications 12:724-734.
Fogarty, M. J., and S. A. Murawski. 1998. Large-scale disturbance and the structure of marine systems: fishery impacts on Georges Bank. Ecological Applications 8:6-22.
Hall, J. S. 1999. The effects of fishing on marine ecosystems and communities. Blackwell Sciences, Oxford, UK.
Hall, M. A. 1998. An ecological view of the tuna-dolphin problem: impacts and trade-offs. Reviews in Fish Biology and Fisheries 8:1-34.
Hampton, J., and D. Fournier. 2001. Stock assessment of yellowfin tuna in the western and central Pacific Ocean. Ocean Fisheries Programme Working Paper SKJ-1, Secretariat of the Pacific Community, Noumea, New Caledonia. Available online at http://www.spc.org.nc/OceanFish/Html/SCTB/SCTB14/skj1.pdf.
Hampton, J., J. R. Sibert, and P. Kleiber. 2003. Comments on Myers & Worm ( Nature, 423:280-283). Available online at http://www.soest.hawaii.edu/PFRP/pdf/Myers_comments.pdf.
Hanamoto, E. 1974. Fishery oceanography of bigeye tuna. I. Depth of capture by tuna longline gear in the eastern tropical Pacific Ocean. La mer 13:58-71.
Hoey, J. J. 1996. Bycatch in Western Atlantic pelagic longline fisheries. Pages 193-203 in Alaska Sea Grant. Solving bycatch: considerations for today and tomorrow. Alaska Sea Grant College Program Report No. 96-03. University of Alaska, Fairbanks, Alaska, USA.
Inter-American Tropical Tuna Commission. 2002a. Resolution on bycatch. Available online at http://www.iattc.org/ PDFFiles/C-02-05%20Bycatch%20resolution%20Jun%2002.pdf.
Inter-American Tropical Tuna Commission. 2002b. The fishery for tunas and billfishes in the eastern Pacific Ocean. Available online at http://www.iattc.org/StockAssessmentReport3ENG.htm.
Ito, R. Y., and W. A. Machado. 2000. Annual report of the Hawaii-based longline fishery for 2000. Southwest Fisheries Science Center Administrative Report H-01-07. Available online at http://www.nmfs.hawaii.edu/fmpi/fmep/hilong/hi2000t.pdf.
Jackson, J. B. C., M. X. Kirby, W. H. Bergner, K. A. Bjorndal, L. W. Botsford, B. J. Bourque, R. H. Bradbury, R. Cooke, J. Erlandson, J. A. Estes, T. P. Hughes, S. Kidwell, C. B. Lange, H. S. Lenihan, J. M. Pandolfi, C. H. Peterson, R. S. Steneck, M. J. Tegner, and R. R. Warner. 2001. Historical overfishing and the recent collapse of coastal ecosystems. Science 293:629-638.
Kitchell, J. F., C. H. Boggs, X. He, and C. J. Walters. 1999. Keystone predators in the central Pacific. Pages 665-704 in Alaska Sea Grant. Ecosystem approaches for fisheries management. University of Alaska, Anchorage, Alaska, USA.
Kitchell, J. F., I. C. Kaplan, S. P. Cox, and C. H. Boggs. 2004. Ecological and economic components of managing rare or endangered species in a tropical pelagic ecosystem. Bulletin of Marine Science 53, in press.
Lee, D. W., and C. J. Brown. 1998. SEFSC Pelagic Observer Program data summary 1992-1996. NOAA Technical Memorandum NMFS-SEFC-408. NOAA Fisheries, Silver Spring, Maryland, USA.
Lewis, A. D., and P. G. Williams. 2001. Overview of the western and central Pacific tuna fisheries. Oceanic Fisheries Programme, Secretariat of the Pacific Community, Noumea, New Caledonia.
Lilly, G. R., D. G. Parsons, and D. W. Kulka. 2000. Was the increase in shrimp biomass on the northeast Newfoundland shelf a consequence of a release in predation pressure from cod? Journal of Northwest Atlantic Fisheries Science 27:45-61.
Link, J. S. 2002. What does ecosystem-based fisheries management mean? Fisheries 27:18-21.
Maunders, M. N., and G. M. Watters. 2001. Status of yellowfin tuna in the eastern tropical Pacific Ocean. Pages 5-86 in Inter-American Tropical Tuna Commission. Stock Assessment Report 1. Inter-American Tropical Tuna Commission, La Jolla, California, USA.
McCoy, M. A., and H. Ishihara. 1999. The socioeconomic importance of sharks in the US flag areas of the western and central Pacific. National Marine Fisheries Service Administrative Report AR-SWR-99-01. National Oceanic and Atmospheric Administration, Washington, D.C., USA.
Myers, R. A., and B. Worm. 2003. Rapid worldwide depletion of predatory fish communities. Nature 423:280-283.
Nakano, H., M. Okazaki, and H. Okamoto. 1997. Analysis of catch depth by species for tuna longline fishery based on catch by branch lines. Bulletin of the National Research Institute of Far Seas Fisheries 34:43-62.
National Archives and Records Administration. 2000. Shark Finning Prohibition Act. (Public Law 106-557). Available online at http://frwebgate.access.gpo.gov/cgi-bin/getdoc.cgi?dbname=106_cong_public_laws&docid=f:publ557.106.
National Marine Fisheries Service. 1999. Ecosystem-based fishery management: a report to Congress by the Ecosystems Principles Advisory Panel. NOAA Fisheries, Silver Spring, Maryland, USA.
National Research Council. 1999. Sustaining marine fisheries. National Academy Press, Washington, D.C., USA.
Olson, R. J., and G. M. Watters. 2003. A model of the pelagic ecosystem in the eastern tropical Pacific Ocean. Inter-American Tropical Tuna Commission Bulletin 22:135-218.
Orange, C. J., and G. C. Broadhead. 1959. 1958–1959—a turning point for tuna purse seine fishing? Pacific Fisherman 57:20-27.
Pauly, D., V. Christensen, J. Dalsgaard, R. Froese, and F. Torres Jr. 1998. Fishing down marine food webs. Science 279:860-863.
Richards, A. H. 1994. Problems of drift-net fisheries in the South Pacific. Marine Pollution Bulletin 29:106-111.
Schindler, D. E., T. E. Essington, J. F. Kitchell, C. Boggs, and R. Hilborn. 2002. Sharks and tunas: fisheries impacts on predators with contrasting life histories. Ecological Applications 12:735-748.
Schmidt, P. G. 1959. The Puretic power block and its effect on modern purse seining. Pages 400-413 in H. Kristjonsson, editor. Modern fishing gear of the world: 3. Fishing News Ltd., London, UK.
Stevens, J. D., R. Bonfil, N. K. Dulvy, and P. A. Walker. 2000. The effects of fishing on sharks, rays, and chimaeras (chondricthyans), and the implications for marine ecosystems. ICES Journal of Marine Sciences 57:476-494.
Tegner, M. J., and P. K. Dayton. 2000. Ecosystem effects of fishing in kelp forest communities. ICES Journal of Marine Sciences 57:579-589.
Walters, C. 1986. Adaptive management of renewable resources. Macmillan, New York, New York, USA.
Walters, C. 2003. Folly and fantasy in the analysis of spatial catch rate data. Canadian Journal of Fisheries and Aquatic Sciences 60:1433-1436.
Walters, C. J., and J. F. Kitchell. 2001. Cultivation/depensation effects on juvenile survival and recruitment: implications for the theory of fishing. Canadian Journal of Fisheries and Aquatic Sciences 58:39-50.
Walters, C. J., D. Pauly, V. Christensen, and J. F. Kitchell. 2000. Representing density dependent consequences of life history strategies in aquatic ecosystems: Ecosim II. Ecosystems 3:70-83.
Watters, G. M., R. J. Olson, R. C. Francis, P. C. Fiedler, J. J. Polovina, S. B. Reilly, K. Y. Aydin, C. H. Boggs, T. E. Essington, C. J. Walters, and J. F. Kitchell. 2003. Physical forcing and the dynamics of the pelagic ecosystem in the eastern tropical Pacific: simulations with ENSO-scale and global-warming climate drivers. Canadian Journal of Fisheries and Aquatic Sciences 60:1161-1175.
Wright, A., and D. J. Doulman. 1991. Drift-net fishing in the South Pacific. Marine Policy 15:303-337.
Yaffe, S. L. 1999. Three faces of ecosystem management. Conservation Biology 13:713-725.
Explanation of animations animation1.gif Animation of the effects of historical fishing effort in the ETP. animation2.gif Animation of the effects of historical fishing effort in the CNP.
Address of Correspondent:
Jefferson T. Hinke
Pacific Fisheries Environmental Laboratory
NOAA Fisheries
1352 Lighthouse Avenue
Pacific Grove, California 93950 USA
Phone: (831) 648-0727
Fax: (831) 648-8440
jhinke@pfeg.noaa.gov
![]()