|
|
|
Copyright © 2004 by the author(s). Published here under license by The Resilience Alliance.
Go to the pdf version of this article.
The following is the established format for referencing this article:
Moen, J., K. Aune, L. Edenius, and A. Angerbjörn. 2004. Potential effects of climate change on treeline position in the Swedish mountains. Ecology and Society 9(1): 16. [online] URL: http://www.ecologyandsociety.org/vol9/iss1/art16/
A version of this article in which text, figures, tables, and appendices are separate files may be found by following this link.
Report Potential Effects of Climate Change on Treeline Position in the Swedish Mountains Jon Moen1, Karin Aune1, Lars Edenius2, and Anders Angerbjörn3
1Department of Ecology and Environmental Science, Umeå University; 2Dept of Animal Ecology, Swedish University of Agricultural Sciences; 3Department of Zoology, Stockholm University
- Abstract
- Introduction
- Methods
- Results
- Discussion
- Responses to this Article
- Acknowledgments
- Literature Cited
Climate change may strongly influence species distribution and, thus, the structure and function of ecosystems. This paper describes simulated changes in the position of the upper treeline in the Swedish mountains in response to predicted climate change. Data on predicted summer temperature changes, the current position of the treeline, and a digital elevation model were used to predict the position of the treeline over a 100-year timeframe. The results show the treeline advancing upward by 233–667 m, depending on the climate scenario used and location within the mountain chain. Such changes hypothetically caused a 75–85% reduction in treeless alpine heaths, with 60–93% of the remaining areas being scree slopes and boulder fields. For this change to occur, the migration rate of the trees would be in the order of 23–221 m yr-1, which is well within published migration rates for wind-dispersed deciduous trees. The remaining alpine areas would be strongly fragmented. These drastic changes would influence all aspects of mountain ecosystems, including biodiversity conservation and human land-use patterns.
KEY WORDS: Betula pubescens ssp. czerepanovii, Climate change, Mountain birch, Treeline dynamics.
Published: May 19, 2004
Climate change is already influencing the distribution of plants and animals globally (IPCC 2001, Parmesan and Yohe 2003, Root et al. 2003). Generally, global meta-analyses have documented significant range shifts toward the poles, or toward higher altitudes, in response to warmer climates, earlier spring conditions, and milder winters (Parmesan and Yohe 2003). These changes may have significant effects on the structure and function of ecosystems (as new species assemblages may form (IPCC 2001)) and, thus, on both conservation values and human utilization patterns. Effects may include changes in primary productivity, extinction of rare species, invasion by more southerly species, and afforestation of previously unforested sites (IPCC 2001). Natural resource managers need to discuss how to adapt to these changes; scenario maps (Peterson et al. 2003) provide a good starting point. Their value lies in facilitating consideration of a variety of possible futures that include many of the uncertainties in the predictions, rather than focusing on a single outcome; they may be used as a basis for developing management strategies for ecosystems. This paper develops such scenario maps for alpine areas in Sweden.
One of the more conspicuous features in alpine areas is the ecotone between forests and treeless alpine heaths. In Fennoscandia, this ecotone is a zone where the continuous birch (Betula pubescens ssp. czerepanovii) forest often gives way rather abruptly to heaths, although there may be individuals or islands of trees above the actual forest line. There is some confusion in the terminology for the various distributional limits of trees but, for the purposes of this paper, we will define “treeline” as the elevation at which continuous forest gives way to alpine heaths and “tree limit” as the elevation of the uppermost individual with a minimum height of 2 m (Kullman 2001). Above this, there is also a “species limit,” which is the highest elevation at which any individual of the species is found.
The position of the treeline is affected by complex interactions with past and current climates and various disturbances (Oksanen et al. 1995, Hofgaard 1997a, 1997b, Körner 1999). Several climatic indices, such as summer mean temperature and temperature sums, are related to treeline position, at least over large scales (Körner 1999, Grace et al. 2002), but it has proved difficult to assign causal factors linking climate and treeline position. It is clear, though, that summer temperatures influence lignification and bud break, and thus growth, in trees (Sveinbjörnsson 2000). Other authors have suggested that the treeline, and the birch forest below the treeline, is in a constant process of recovery from previous disturbances (e.g., Emanuelsson 1987, Hofgaard 1999), which would make causal inferences with climate hard to detect.
One important disturbance factor influencing growth at the treeline is insect defoliation, especially from outbreaks of geometrid moths (Tenow et al. 2001). Such outbreaks may result in the retreat of the treeline over vast areas, if the defoliation causes mortality of established trees (Neuvonen et al. 2001). Mortality risk is also related to the amount of stored resources in the trees, and interactions with reindeer (Rangifer tarandus) grazing on resprouting shoots (Lehtonen and Heikkinen 1995, Neuvonen et al. 2001). In some places, human land use, such as livestock grazing and firewood gathering, is also an important factor influencing forest structure at, or close to, the treeline (Hofgaard 1999). The birch forest and treeline have also been continuously grazed by reindeer, probably since the retreat of the last ice age about 8000-10 000 years BP. Birch leaves are a preferred summer food for reindeer (Helle 2001) and grazing can alter the structure of treeline forests from brushwood to savanna-like stands (Oksanen et al. 1995).
The position of the tree limit has been suggested as a sensitive indicator of climate change (Kullman 1998), because an improved climate would be expected to rapidly influence growth of established individuals. It is less clear whether the position of the treeline would respond as quickly as the tree limit to a changed climate as this would involve dieback of trees or establishment of new individuals. Nevertheless, treeline position has also been suggested as an indicator of climate change (Holten and Carey 1992). What is clear, however, is that climate change has the potential to affect the position of the treeline. This will have consequences for land use in the region, especially for reindeer husbandry and tourism, and may also have long-term consequences for biodiversity conservation. For example, increased afforestation will increase the probability of insect attack on the reindeer, with consequent negative effects on animal growth (Danell et al. 1999, Hagemoen and Reimers 2002). Furthermore, a decrease in the area of open heath will reduce recreational and aesthetical values with negative effects on the summer tourist industry in the Swedish mountain region, and a reduced and fragmented alpine zone will probably increase the extinction rates of alpine species (Bruun and Moen 2003).
This study aims to simulate changes in the position of the alpine treeline in Sweden in response to predicted changes in climate over a 100-year timeframe. This will provide an indication of the scale of a worst-case scenario where climate change overrides other factors in determining treeline position. Given our current level of knowledge. we cannot produce a dynamic, mechanistic model of tree growth at the scale of the entire mountain chain, but we can create a static simulation of potential treeline changes as a scenario-planning tool. These scenario maps may be used as a basis for discussions with scientists and stakeholders on how best to manage alpine areas and conserve biodiversity in the face of climate change.
For this analysis, we assume that the current position of the birch treeline in the Swedish mountain region (Fig. 1) is determined by, and in balance with, climatic factors, and more specifically the mean summer (June–August) temperature. Summer temperature was chosen as it correlates with treeline position over large scales (Körner 1999, Grace et al. 2002). Furthermore, we assume that the trees have a migration rate that is fast enough to track changes in summer temperature, at least within the scale of the analysis. This implies that a change in mean summer temperature will produce a change in the position of the treeline. We convert changes in temperature to changes in elevation by using a lapse rate of 0.6°C/100 m. This lapse rate is typical for the area (Swedish National Atlas 1995). It is possible that the lapse rate varies over both space and time, but we have no data from our study area that allow us to vary this parameter. This means that, for example, an increase of 1.2°C will give a 200-m rise in the treeline. Similar reasoning has been used when inferring past temperatures from pollen and macrofossil finds of trees (e.g., Barnekow and Sandgren 2001, Seppä and Birks 2001, 2002), and may be considered the simplest way to describe correlations between climate and treelines without incorporating mechanistic details.
|
Fig. 1. Map of Sweden, showing the counties mentioned in the paper. The hatched area indicates the mountain region and the yellow square indicates the delimited area shown in greater detail in Fig. 6.
|
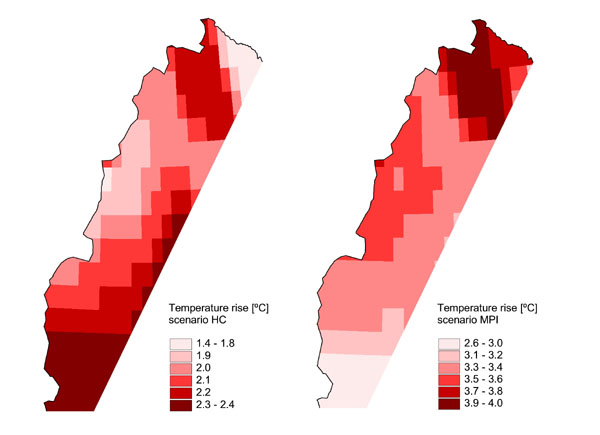
Data on predicted changes in summer temperature (mean values for June, July, and August) were provided by SweClim, a Swedish regional climate-modeling project. SweClim has modeled the regional climate of Fennoscandia based on two different general circulation models, one from the Hadley Centre, UK, and one from the Max-Planck Institut, Germany (hereafter called scenarios HC and MPI, respectively; see Rummukainen et al. (2001) and Räisanen et al. (2001, 2004) for details on the regional climate models). In short, the climate data provided for this paper were modeled at a 0.5-degree grid cell resolution (20–27 x 56 km) and consisted of the predicted values after 100 years of simulation, i.e., roughly at year 2100 (Fig. 2). The two scenarios give different predictions for the increases in summer temperature, with scenario HC predicting increases of 1.4–2.4°C, and scenario MPI giving predictions of 2.6–4.0°C. The difference between the scenarios reflects differences in the assumptions and structure of the general circulation models. However, both models predict the same magnitude of global temperature increase (Räisanen et al. 2001) and are considered to encompass the current predictions of climate change patterns in the region (Rummukainen et al. 2001, Räisanen et al. 2004).
|
Fig. 2. Predicted increases in mean summer temperature (June–August) based on general circulation models from the Hadley Centre (scenario HC) on the left, and from the Max-Planck Institute (scenario MPI) on the right. (Data from SweClim, SMHI, Sweden.)
|
Geographical information systems (GIS) were used to simulate treeline changes. We used digital maps covering the entire Swedish mountain area (ca. 118 000 km2) to determine the position of the current treeline. Topography in the area was described using a digital elevation model (DEM) with a resolution of 50 x 50 m pixels. This pixel size is also the scale of the simulation. The climate predictions (converted to elevation using the lapse rate) were transformed into the same pixel size and extension as the DEM and formed a new climate grid. Pixels in this grid originating from the same climate grid cell (0.5° cell size) obtained the same value. We were thus unable to take local, small-scale variations in climate into account. However, aspect is incorporated in the sense that the current treeline position (and thus the starting point for the simulation) is lower on the north-facing slopes of the mountains.
The location of the current treeline (described as polylines), the DEM, and the climate grid were used as input data in the model. A script in ArcView® (ver. 3.2a) was programmed to calculate new locations for the treeline depending on the climate scenarios. The mountain region was divided into 47 million pixels of 50 x 50 m. For each pixel, the script found the closest treeline of today and, depending on the climate grid, the altitude of the current treeline, and the altitude of the pixel, determined whether the pixel became forested or not. We incorporated the added conservative assumption that trees would only grow on sites that currently had soil cover, as soil formation is a slow process in the mountains. Therefore, we used digital vegetation maps to exclude any area containing only gravel, rocks, or glaciers from the predicted area covered by forests. In addition to the two climate scenarios, we included a more conservative scenario of a 100-m treeline increase (hereafter referred to scenario 100 m) in order to get some information on early changes as the treeline advances. This scenario was treated in the same way as the two climate scenarios. Furthermore, we calculated the area covered by treeless alpine heaths in the different scenarios, and the fragmentation of the alpine area.
We also calculated the mean, median, and maximum horizontal distance that the trees would have to migrate under the most extreme climate scenario (scenario MPI). We randomly chose 4000 points over the entire treeline calculated according to scenario MPI, and for each of these points we measured the nearest distance to the current treeline.
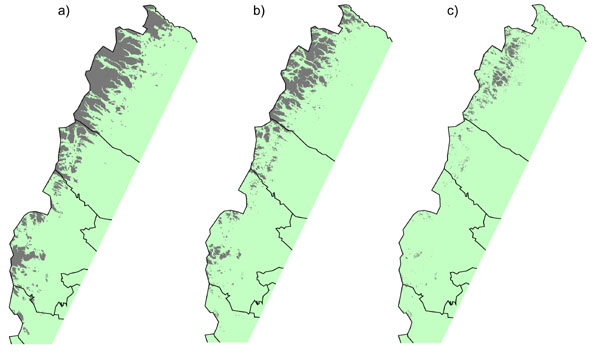
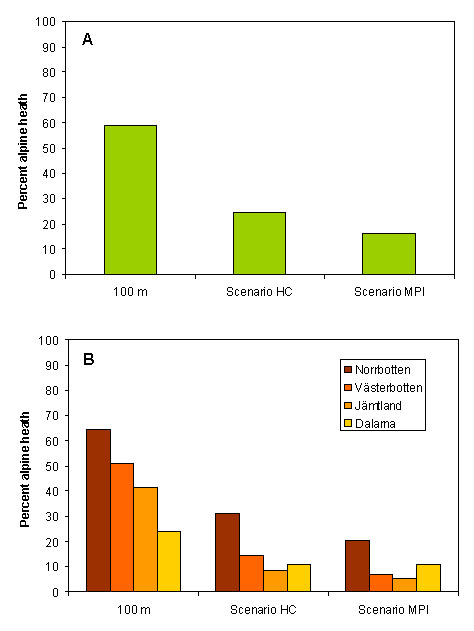
The analysis shows a rise in treeline position of 233–667 m, depending on the climate scenario and on location in the mountain chain (Fig. 3). According to scenario MPI, only very small alpine areas would remain, and most of these would be in the northern part of the mountain chain, where topography is steeper and the mountains reach higher altitudes. Thus, there would be a substantial reduction in treeless alpine heaths (by 75% according to scenario HC and by 85% according to scenario MPI) (Fig. 4, Table 1). The more conservative estimate of a 100-m rise in the treeline shows that even this relatively moderate advance would reduce alpine heath area by 41%. The results further show that much of the remaining alpine areas would be composed of scree slopes and boulder fields (100 m: 25%, scenario HC: 60%, and scenario MPI: 93%).
|
Fig. 3. Current and simulated distributions of treeless alpine heaths in Sweden (areas marked in gray): a) current distribution; b) simulated distribution assuming a 100-m altitudinal advance of the treeline; and c) simulated distribution according to climate scenario MPI (predicted increase of the treeline: 433–667 m).
|
|
Fig. 4. Predicted area of treeless alpine heaths as a percentage of the current distribution for A) the whole of Sweden, and B) each county.
|
The trees would have to migrate on average 2.3 km under the most extreme climate scenario (MPI: median distance–1.3 km, maximum distance–22.1 km). This gives an average migration rate of 23 m yr-1, and a maximum migration rate of 221 m yr-1, which is well within published migration rates of wind-dispersed deciduous trees (Huntley 1991, Wilkinson 1997).
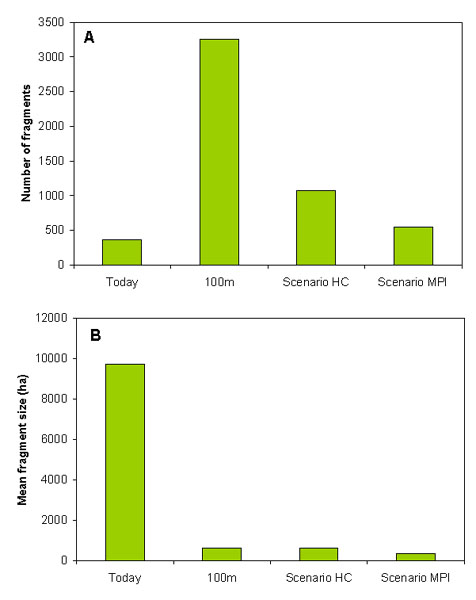
Advances in the treeline would also fragment the remaining alpine heaths. Currently, alpine heaths in the Swedish mountain region are divided into 362 areas with a mean size of 9700 ha (Fig. 5). The smallest areas occur in the southern part of the mountain chain and in some isolated eastern outposts. However, the northern part of the mountain chain consists of large expanses of alpine heaths. A moderate advance in the treeline of 100 m would result in an increase in alpine heath fragments by an order of magnitude (Fig. 5). This is because lower lying valleys, that today are treeless, would be covered by forests, which would break up the large treeless areas into many smaller areas. As the treeline advances even further (as suggested by the climate scenarios), the number of fragments would quickly decline as low altitude mountains become completely covered by forests. Under the most extreme climate scenario, the remaining fragments would, on average, be only about 4% of the current average size.
|
Fig. 5. Predicted fragmentation of treeless alpine areas: A) number of fragments; B) mean size of fragments.
|
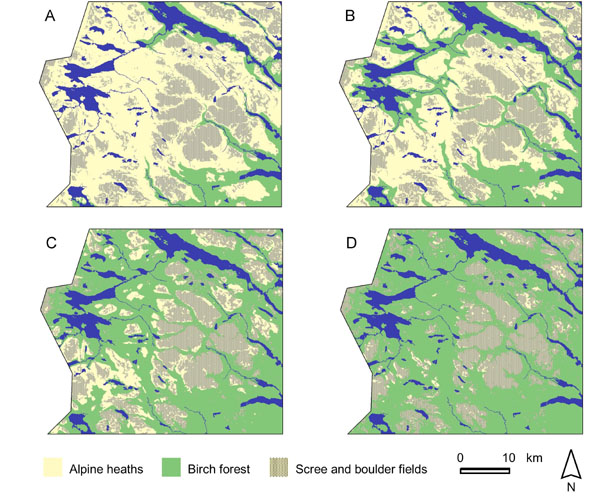
An example from the largest national park complex in Sweden (the Sarek-Padjelanta area) illustrates these effects (Fig. 6). A 100-m advance of the treeline would cause the birch forest to invade the valleys on both the east and west side of the area, reducing alpine heath area by 19%. According to scenario HC and, even more extremely, scenario MPI, most of the western part would be covered by birch forests, reducing alpine heath area by 53 and 67%, respectively. However, most of the remaining alpine areas would be scree slopes and boulder fields as the high peaks in Sarek are very steep and have sparse vegetation.
|
Fig. 6. Example of simulated increases in treeline position from the Sarek area, northern Sweden. A. The current situation. B. After a 100-m altitudinal advance of the treeline (distribution of treeless alpine heaths: 81% of the current area). C. According to climate scenario HC (alpine heaths: 47% of the current area). D. According to scenario MPI (alpine heaths: 33% of the current area).
|
An advancement in treeline position, as suggested in this study, would completely change the appearance of the mountain region. There would probably also be other changes in the distribution of alpine species, which in turn would change the structure and function of the ecosystem (e.g., Walker and Steffen 1997). For example, it is likely that species composition will change as different species respond differently to changes in climate. This will have effects on both biodiversity patterns and land use, such as reindeer husbandry, in the area.
There is already a general trend toward increased growth in high altitude areas. Several reports show an increased growth of established trees and saplings above the treeline, which, in places, has increased the tree limit by ca. 100 m (Kjällgren and Kullman 1998, Kullman 2000, 2002b). Fast-growing saplings have also occurred more than 300 m above the tree limit (Kullman 2002b), and these advances are attributed to a series of warm summers and mild winters (Kullman 2001). This may be a general phenomenon in sub-Arctic and Arctic sites, as a widespread increase in shrub abundance, covering more than 320 km2, has been shown from Alaska (Sturm et al. 2001). Global meta-analyses have also documented significant range shifts toward the poles or toward higher altitudes for many organisms, and a large part of these changes may be attributed to increased global temperatures (Parmesan and Yohe 2003, Root et al. 2003).
The simulated treeline advancement (in the order of 233–667 m) is not unreasonable from a historical point of view. Birch macrofossils have been found 300–400 m above the current treeline in Abisko, Sweden (Barnekow and Sandgren 2001), suggesting that birch forests covered the area in the early (ca. 9000 years BP) and middle (ca. 5000 years BP) Holocene. Similarly, Kullman (1993) found subfossils of birch, dated to 4000–5000 BP, at locations 200–300 m above the current tree limit in Västerbotten, Sweden, and 300–500 m above the current tree limit at Mt. Åreskutan, Jämtland (dated to 5000–14 000 BP; Kullman 2002a). This suggests that birch forests exhibit long-term transitions up and down the mountain in response to long-term climatic changes. The large expanses of treeless alpine heaths that we see today may thus be seen as an interstadial between more forested time periods.
The advance in the treeline will fragment the current large continuous expanses of alpine heaths. This fragmentation may increase risk of species extinction as populations get smaller and dispersal distances increase. Climate change will force many alpine species toward higher altitudes, both as a result of afforestation and a changed climate. An overall trend of upward migrations of alpine species on mountain summits in the Alps has already been detected (Grabherr et al. 1994).
When calculating predicted changes, there are of course large uncertainties inherent in the climate modeling, which we presently cannot improve. However, all models are certain that warming will occur, so the uncertainties lie in how much and how fast warming will proceed, and in the spatial variations. It is important to note that we only considered mean changes in temperature. It is likely that climate extremes will increase, such as maximum or minimum temperatures or precipitation (Räisanen and Joelsson 2001), which may affect tree growth and thus treeline dynamics. For example, if situations with very low temperatures increase in frequency, the treeline advance may slow down because of tree and bud mortality.
We also need to scrutinize the importance of our assumptions for calculating these changes in treeline position. We assumed that the treeline was correlated with summer climate. Although it is clear that climate plays an important role in determining the treeline (e.g., Körner 1998, 1999, Grace et al. 2002), other processes are also important, such as soil conditions, winter dessication, seed limitation, and competition from established field layer vegetation (cf. Dullinger et al. 2004). These factors, together with the carbon balance of the established trees, should be included in a dynamic model of tree growth at treelines (Cairns and Malanson 1998).
Ellenberg (1988) discussed two types of treelines, transitional and abrupt. He argued that, in completely climatic-driven treelines, a transitional treeline would appear with progressively smaller trees as climate became more unfavorable, until eventually the species could only survive as stunted or krummholz individuals. However, when herbivores are present, browsing would deplete resources from individuals already living on the margin and thus increase mortality. This would result in an abrupt treeline consisting of established trees that have extended beyond the browsing line in times of relaxed grazing pressure. Browsing would also prevent regeneration and very few saplings and young trees would be found.
In Fennoscandia, the treeline is grazed by semi-domesticated reindeer and grazing may affect both the position and the structure of the treeline (Oksanen et al. 1995, Hofgaard 1997b). In areas with summer grazing, the treeline is typically abrupt and consists of large trees, whereas the treeline is more transitional where grazing is less intensive. Birch saplings may be found several hundred meters above treeline, but they are generally browsed and resemble very small coppiced trees. Studies in Finland have shown that reindeer may consume up to 90% of the total birch leaf biomass available in the 0–130 cm layer above the ground (Helle 2001). Thus, it seems likely that grazers have the ability to slow, or even halt, the upward spread of the treeline by feeding on seedlings and saplings already weakened by adverse climatic conditions and competition from field layer plants. This warrants further studies in order to incorporate herbivory in future models on treeline responses to climate change.
We also assumed that the birch trees would be able to migrate fast enough to respond to changes in climate. We do not have any direct information with which to verify this, however, there is some indirect evidence supporting this assumption. As already mentioned, single birch trees and saplings are commonly found above the treeline, and birch seeds have wings that facilitate wind dispersal. Wind-dispersed deciduous trees typically have migration rates in the order of 150–750 m yr-1 (Huntley 1991, Wilkinson 1997), which is well within the simulated migration rates in this study (average rate: 23 m yr-1, maximum rate: 221 m yr-1). Furthermore, Molau and Larsson (2000) found seeds of mountain birch both in the seed rain and in the seed bank at a site 300 m above, and some 5–8 km away from, the current treeline (Abisko, northern Sweden), which illustrates that seed dispersal within the calculated range occurs today.
In conclusion, climate change will likely cause the current birch treeline to advance upward. Our analysis shows altitudinal increases in the order of 233–667 m within the next century. This will result in a reduction of 75–85% in the current treeless alpine areas, and 60–93% of the remaining areas will be scree slopes and boulder fields. This drastic change will influence all aspects of mountain ecosystems, including biodiversity patterns and the potential for reindeer husbandry and tourism development.
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a comment, follow this link. To read comments already accepted, follow this link.
We thank Marko Rummukainen at SweClim for the climate prediction data, and David Cairns and two anonymous reviewers for comments on the paper. This project was financed by MISTRA through the Mountain Mistra Programme.
Barnekow, L., and P. Sandgren. 2001. Palaeoclimate and tree-line changes during the Holocene based on pollen and plant macrofossil records from six lakes at different altitudes in northern Sweden. Review of Palaeobotany and Palynology 117:109–118.
Bruun, H. H., and J. Moen. 2003. Nested communities of alpine plants on isolated mountains: relative importance of colonization and extinction. Journal of Biogeography 30:297–303.
Cairns, D. M., and G. P. Malanson. 1998. Environmental variables influencing carbon balance at the alpine treeline ecotone: a modeling approach. Journal of Vegetation Science 8:679–692.
Danell, Ö., Ø. Holand, H. Staaland, and M. Nieminen. 1999. Reindeer adaptations and nutrient requirements. (In Swedish.) Pages 31–46 in H. K. Dahle, Ö. Danell, E. Gaare, and M. Nieminen, editors. Reindrift in Nordvest-Europa i 1998—biologiske muligheter og begrensninger. TemaNord 1999:510.
Dullinger, S., T, Dirnböck, and G. Grabherr. 2004. Modelling climate change-driven treeline shifts: relative effects of temperature increase, dispersal and invasibility. Journal of Ecology 92:241–252.
Ellenberg, H. 1988. Vegetation ecology of Central Europe. 4th ed. Cambridge University Press, Cambridge, UK.
Emanuelsson, U. 1987. Human influence on vegetation in the Torneträsk area during the last three centuries. Ecological Bulletin 38:95–111.
Grabherr, , G., M. Gottfried, and H. Pauli. 1994. Climate effects on mountain plants. Nature 369:448.
Grace, J., F. Berninger, and L. Nagy. 2002. Impacts of climate change on the tree line. Annals of Botany 90:537–544.
Hagemoen, R. I. M., and E. Reimers. 2002. Reindeer summer activity pattern in relation to weather and insect harassment. Journal of Animal Ecology 71:883–892.
Helle, T. 2001. Mountain birch forests and reindeer husbandry. Pages 279–291 in F. E. Wielgolaski, editor. Nordic mountain birch ecosystems. Man and the Biosphere Series, vol. 27, Parthenon Publishing, New York, New York, USA.
Hofgaard, A. 1997a. Inter-relationships between treeline position, species diversity, land use and climate change in the central Scandes mountains of Norway. Global Ecology and Biogeography Letters 6:419–429.
Hofgaard, A. 1997b. Structural changes in the forest–tundra ecotone: a dynamic process. Pages 255–263 in B. Huntley, W. Cramer, A. V. Morgan, H. C. Prentice, and J. R. M. Allen, editors. Past and future rapid environmental changes. Springer, Berlin, Germany.
Hofgaard, A. 1999. The role of “natural” landscapes influenced by man in predicting responses to climate change. Ecological Bulletin 47:160–167.
Holten, J. I., and P. D. Carey. 1992. Responses of climate change on natural terrestrial ecosystems in Norway. NINA Research Report 29:1–59.
Huntley, B. 1991. How plants respond to climate change: migration rates, individualism and the consequences for plant communities. Annals of Botany 67 (Suppl. 1):15–22.
IPCC (Intergovernmental Panel on Climate Change). 2001. Climate change 2001. Impacts, adaptation, and vulnerability. Cambridge University Press, Cambridge, UK.
Kjällgren, L., and L. Kullman. 1998. Spatial patterns and structure of the mountain birch tree-limit in the southern Swedish Scandes – a regional perspective. Geografiska Annaler 80A:1–16.
Körner, C. 1998. A re-assessment of high elevation treeline positions and their explanations. Oecologia 115:445–459.
Körner, C. 1999. Alpine plant life. Functional plant ecology of high mountain ecosystems. Springer, Berlin, Germany.
Kullman, L. 1993. Holocene thermal trend inferred from tree-limit history in the Scandes mountains. Global Ecology and Biogeography Letters 2:181–188.
Kullman, L. 1998. Tree-limits and montane forests in the Swedish Scandes: sensitive biomonitors of climate change and variability. Ambio 27:312–321.
Kullman, L. 2000. Tree-limit rise and recent warming: a geoecological case study from the Swedish Scandes. Norsk Geografisk Tidskrift 54:49–59.
Kullman, L. 2001. 20th century climate warming and tree-limit rise in the southern Scandes in Sweden. Ambio 30:72–80.
Kullman, L. 2002a. Boreal tree taxa in the central Scandes during the late-glacial: implication for late-Quaternary forest history. Journal of Biogeography 29:1117–1124.
Kullman, L. 2002b. Recent reversal of Neoglacial climate cooling trend in the Swedish Scandes as evidenced by mountain birch tree-limit rise. Global Planetary Change 36:77–78.
Lehtonen, J., and R. K. Heikkinen. 1995. On the recovery of mountain birch after Epirrita damage in Finnish Lapland, with a particular emphasis on reindeer grazing. Ecoscience 2:349–356.
Molau, U., and E.-L. Larsson. 2000. Seed rain and seed bank along an alpine altitudinal gradient in Swedish Lapland. Canadian Journal of Botany 78:728–747.
Neuvonen, S., K. Ruohomäki, H. Bylund, and P. Kaitaniemi. 2001. Insect herbivores and herbivory effects on mountain birch dynamics. Pages 279–291 in F. E. Wielgolaski, editor. Nordic mountain birch ecosystems. Man and the Biosphere Series, vol. 27. Parthenon Publishing, New York, New York, USA.
Oksanen, L., J. Moen, and T. Helle. 1995. Timberline patterns in northernmost Fennoscandia. Relative importance of climate and grazing. Acta Botanica Fennica 153:93–105.
Parmesan, C., and G. Yohe. 2003. A globally coherent fingerprint of climate change impacts across natural systems. Nature 421:37–42.
Peterson, G. D., G. S. Cumming, and S. R. Carpenter. 2003. Scenario planning: a tool for conservation in an uncertain world. Conservation Biology 17:358–366.
Räisanen, J., and R. Joelsson. 2001. Changes in average and extreme precipitation in two regional climate model experiments. Tellus A 53:547–566.
Räisanen, J., M. Rummukainen, and A. Ullerstig. 2001. Downscaling of greenhouse gas induced climate change in the GCMs with the Rossby Centre regional climate model for northern Europe. Tellus A 53:168–191.
Räisanen, J., U. Hansson, A. Ullerstig, R. Döscher, L. P. Graham, C. Jones, H. E. M. Meier, P. Samuelsson, and U. Willén. 2004. European climate in the late twenty-first century: regional simulations with two driving global models and two forcing scenarios. Climate Dynamics 22:13–31.
Root, T., J. T. Price, K. R. Hall, S. H. Schneider, C. Rosenzweig, and J. A. Pounds. 2003. Fingerprints of global warming on wild animals and plants. Nature 421:57–60.
Rummukainen, M., J. Räisanen, B. Bringfelt, A. Ullerstig, A. Omstedt, U. Willén, U. Hansson, and C. Jones. 2001. A regional climate model for northern Europe: model description and results from the downscaling of the GCM control simulations. Climate Dynamics 17:339–359.
Seppä, H., and H. J. B. Birks. 2001. July mean temperature and annual precipitation trends during the Holocene in the Fennoscandian tree-line area: pollen-based climate reconstructions. The Holocene 11:527–539.
Seppä, H., and H. J. B. Birks. 2002. Holocene climate reconstructions from the fennoscandian tree-line area based on pollen data from Toskaijavri. Quartenary Research 57:191–199.
Sturm, M., C. Racine, and K. Tape. 2001. Increasing shrub abundance in the Arctic. Nature 411:546–547.
Sveinbjörnsson, B. 2000. North American and European treelines: external forces and internal processes controlling position. Ambio 29:388–395.
Swedish National Atlas. 1995. Climate, lakes and rivers. Lantmäteriet, Gävle, Sweden.
Tenow, O., H. Bylund, and B. Holmgren. 2001. Impact on mountain birch forests in the past and the future of outbreaks of two geometrid moths. Pages 223–239 in F. E. Wielgolaski, editor. Nordic mountain birch ecosystems. Man and the Biosphere Series, vol. 27. Parthenon Publishing, New York, New York, USA.
Walker, B., and W. Steffen. 1997. An overview of the implications of global change for natural and managed terrestrial ecosystems. Conservation Ecology: 1(2):2. (online.) URL: http://www.ecologyandsociety.org/vol1/iss2/art2.
Wilkinson, D. M. 1997. Plant colonization: are wind dispersed seed really dispersed by birds at larger spatial and temporal scales? Journal of Biogeography 24:61–65.
Address of Correspondent:
Jon Moen
Department of Ecology and Environmental Science,
Umeå University,
SE-901 87 Umeå
Sweden
Phone: +46-90-7869647
Fax: +46-90-7866705
jon.moen@eg.umu.se
![]()