|
|
|
Copyright © 2004 by the author(s). Published here under license by The Resilience Alliance.
Go to the pdf version of this article.
The following is the established format for referencing this article:
Whittington, J., C. C. St. Clair, and G. Mercer. 2004. Path tortuosity and the permeability of roads and trails to wolf movement. Ecology and Society 9(1): 4. [online] URL: http://www.ecologyandsociety.org/vol9/iss1/art4/
A version of this article in which text, figures, tables, and appendices are separate files may be found by following this link.
Report Path Tortuosity and the Permeability of Roads and Trails to Wolf Movement Jesse Whittington1, Colleen Cassady St. Clair1, and George Mercer2
1University of Alberta; 2Jasper National Park
- Abstract
- Introduction
- Methods
- Results
- Discussion
- Responses to this Article
- Acknowledgments
- Literature Cited
Few studies have examined the effects of human development on fine-scale movement behavior, yet understanding animal movement through increasingly human-dominated landscapes is essential for the persistence of many wild populations, especially wary species. In mountainous areas, roads and trails may be particularly deserving of study because they are concentrated in the valley bottoms where they can impede animal movement both across and between valleys. In this study, we tracked wolf (Canis lupus) movement in the snow for two winters in Jasper National Park, Alberta, Canada to examine how wolves navigate through or around human-use features. We quantified the effects of human development and topography on the tortuosity of wolf paths and then tested the permeability of roads, trails, and a railway line to wolf movement by comparing the frequency with which actual wolf paths and a null model of random paths crossed these features. Wolf path tortuosity increased near high-use trails, within areas of high-trail and road density, near predation sites, and in rugged terrain. Wolves crossed all roads, trails, and the railway line 9.7% less often than expected, but avoided crossing high-use roads more than low-use trails. Surprisingly, trails affected movement behavior of wolves equally, if not more, than roads. These results suggest that although roads and trails in this study were not absolute barriers to wolf movement, they altered wolf movements across their territories.
Published: January 21, 2004
Human developments create immense problems for wildlife the world over because they degrade habitat quality. Of the many types of human developments, roads are one of the most well-studied and problematic sources of habitat degradation and fragmentation (Forman 2000, Trombulak and Frissell 2000). Although roads are known to degrade habitat quality for many organisms (Mace et al. 1996, Mladenoff et al. 1999, Rowland et al. 2000, Dyer et al. 2001), few studies have quantified the effects of roads and other human developments on fine-scale movement behavior (e.g., Bélisle and St. Clair 2001, Rondinini and Doncaster 2002). Such detailed emphasis on movement behavior may identify sources of disturbance that are not identifiable with traditional habitat-use studies (Caro 1998, Desrochers and Fortin 2000). The ability of animals to move through poor-quality habitats or places where human activity is concentrated may determine whether or not habitat fragmentation constrains the kind of movement, such as dispersal, that most affects population viability (With and Crist 1995, Keitt et al. 1997, Schultz 1998, Turchin 1998, Brooker et al. 1999, With and King 1999, Roland et al. 2001, Brooker and Brooker 2002).
Studies linking fine-scale movement behavior to habitat quality have generally measured movement behavior either in terms of path complexity (tortuosity) or propensity to travel across inhospitable habitats. Tortuosity is a useful metric because it may be expected to reflect habitat quality (Odendaal et al. 1989, Crist et al. 1992, Miyatake et al. 1995, Stapp and Van Horne 1997, Etzenhouser et al. 1998, Gillis and Nams 1998, Schultz 1998, Kindvall 1999, Schultz and Crone 2001) and energy expenditure (Wiens et al. 1995). Several non-anthropogenic factors appear to influence path tortuosity, including stage of life history, habitat complexity, and habitat quality. For example, path tortuosity may change seasonally when species are migrating (Bergman et al. 2000) or denning (Bascompte and Vilá 1997). In terms of habitat complexity, some insects with a clumped distribution of resources maximize foraging efficiency by decreasing the tortuosity of their paths (Wiens et al. 1995). Similarly, when some species are within high-quality habitat, they maximize foraging efficiency by decreasing their speed of travel and increasing the tortuosity of their paths (Odendaal et al. 1989, Crist et al. 1992, Miyatake et al. 1995, Stapp and Van Horne 1997, Etzenhouser et al. 1998, Gillis and Nams 1998, Schultz 1998, Kindvall 1999, Schultz and Crone 2001). Although none of these studies has directly assessed the influence of roads, trails, or other developments on path tortuosity, the response of several species to habitat quality suggests that where human developments degrade habitat quality, lower tortuosity may be expected.
In addition to understanding how organisms change their movement behavior near human developments, the distribution of a species in anthropogenically altered landscapes may also depend on their ability to cross roads, trails, and other linear features. Several authors have successfully manipulated birds, inducing them to cross anthropogenic barriers using audio playbacks (e.g., Desrochers and Hannon 1997, St. Clair et al. 1998, St. Clair 2003, in press) or translocating territorial individuals (e.g., Bélisle et al. 2001, Gobeil and Villard 2002). These studies have demonstrated that some bird species typically avoid crossing inhospitable habitat at both small (e.g, 50-m) and larger (e.g., 2-km) scales. The configuration of barriers may also be important (Bélisle and St. Clair 2001, Brooker and Brooker 2002) and is sometimes apparent even in observational studies (e.g., Brooker et al. 1999, Desrochers and Fortin 2000). Translocations of forest-dependent mammals suggest that the ability of some microtines and sciurids to cross inhospitable habitats depends on their perceptual range (Zollner and Lima 1997, Gillis and Nams 1998, Zollner 2000; but see Bowman and Fahrig 2002). As an alternative to playback and relocation experiments, other studies of mammal movement compared the frequency with which animals cross roads to a null model of barrier crossings. Such studies found that black bears (Ursus americanus), caribou (Rangifer tarandus), and hedgehogs (Erinaceus europaeus) crossed high-use roads less often than expected, whereas low-use roads and seismic lines did not seem to affect their movement patterns (Serrouya 1999, Dyer 1999, Rondinini and Doncaster 2002, respectively). In these studies, the animals were assumed to move in a correlated or uncorrelated random walk. In the present study, we expand upon this approach by first testing the appropriateness of three random walk models for wolf (Canis lupus) movement and then using the best model to create a null model of barrier crossings.
The effects of human development on animal movement are accentuated in mountainous areas. Here, rugged topography and deep snows confine the movement of animals to the valley bottoms where people also concentrate their activity (Noss et al. 1996). Therefore, there is high potential for human activity to obstruct animal movement across or between valleys (Clevenger and Waltho 2000, Bélisle and St. Clair 2001). The wolf is a wary species that may be especially susceptible to these habitat fragmentation effects (Weaver et al. 1996, Mladenoff et al. 1999). Wolves often travel > 30 km in a day within territories that encompass several valleys and ~1000 km2. Consequently, wolf movement through areas with human activity is often necessary for their local persistence.
In this study, we address the effects of human development on the fine-scale movement of wolves around the mountainous town of Jasper, Alberta, Canada. We examine the effects of human development on wolf movement behavior in two ways. We first determine how roads, trails, railway lines, and commercial accommodations affect the tortuosity of wolf paths. At the same time, we account for other factors, such as terrain ruggedness and proximity to predation sites, that may also influence path tortuosity. We then test whether high- and low-use roads, trails, and a railway line present barriers to wolf movement. For this analysis, we compare the frequency with which wolf paths and a null model of simulated wolf paths cross these features.
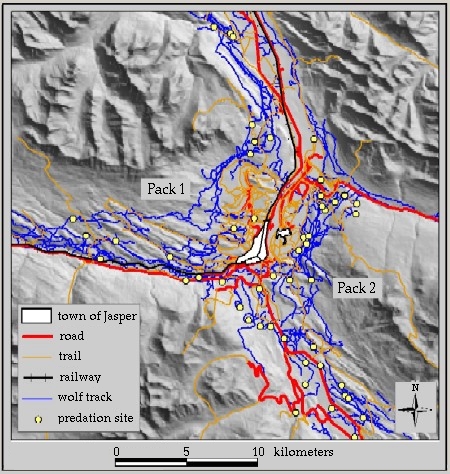
This study focused on the movement of wolves within 20 km of the hamlet of Jasper within Jasper National Park, Alberta, Canada (52o 52’ N, 188 o 05’ W, elevation 1040–2800 m). Jasper National Park lies along the eastern side of the Rocky Mountains, where snow depths along the valley bottoms are generally shallow and range from 5 cm to 40 cm. However, deeper snows and rugged topography found at higher elevations confine the winter movement of wolves to three valleys that converge upon the town site of Jasper. Two wolf packs traveled through the study area during the study. Pack 1, locally referred to as the Decoigne Pack, consisted of 7–10 wolves that ranged west and northeast of Jasper. Pack 2, locally referred to as the South Pack, consisted of 2–3 wolves that ranged south and southeast of Jasper. These wolves preyed upon elk (Cervus elephus), deer (Odocoileus spp.), moose (Alces alces), and occasionally big-horned sheep (Ovis canadensis, caribou (Rangifer tarandus), and mountain goat (Oreamnos americanus).
The study area included 262 km of roads, 422 km of trails below 1500 m, and a railway line. A major highway through the study area follows valleys from northeast to west of the Jasper town site and is neither divided nor fenced. However, it received 1,288,788 vehicles in 2000, including substantial freight truck traffic (Parks Canada Highway Services, unpublished data). Vehicle traffic quadruples from winter to summer and has increased by 22% in the last decade. Several secondary highways also extend throughout Jasper National Park. Human use on trails increases ~20fold from winter to summer (Parks Canada, unpublished data). Trail networks are concentrated within 10 km of the Jasper town site, but are rapidly expanding as people, particularly mountain bikers, create their own trails throughout lower elevations in the study area.
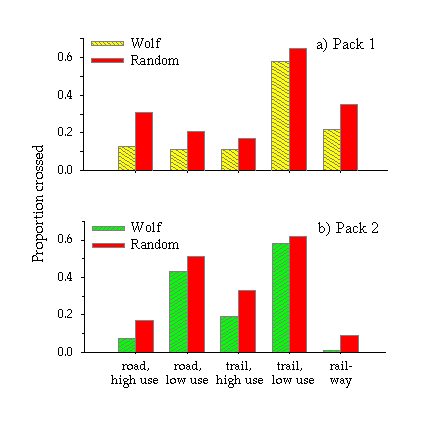
Roads and trails were divided into two categories: high- or low-use. In February, high-use roads were estimated to receive > 10,000 vehicles per month, while low-use roads were estimated to receive < 10,000 vehicles per month and had more pronounced diurnal variation in traffic volume. High-use trails received foot traffic on a daily basis in winter, whereas low-use trails received infrequent or no foot traffic. The railway line received ~30 freight and passenger trains each day. When comparing the length of linear features encompassed by each pack’s range of movement within the study area, Pack 1 had more high-use roads (59 km vs. 31 km), fewer low-use roads (41 km vs. 131km), fewer high-use trails (42 km vs. 60 km), and an equal number of low-use trails (~160 km each). The railway line occurred mostly within Pack 1’s territory.
Field methodsWe snow-tracked the movements of the two wolf packs for two winters (1999–2000, 2000–2001). Wolf tracks were initially located by conducting road surveys and cross-valley transects. Once found, wolf tracks were then followed in the direction opposite to wolf travel (i.e., backtracking) while snow and light conditions permitted. While tracking, we automatically recorded wolf locations every 25 m with a hand-held global positioning system (GPS; Trimble GeoExplorer3; Trimble, Sunnyvale, California, USA). Before selective availability was turned off (1 May 2000), all wolf locations were differentially corrected. Consequently, we considered wolf locations to be accurate within 25 m. Wolves usually traveled in single file, but when wolf tracks diverged, we followed the path made by the majority of wolves. While tracking, we also recorded what the wolves traveled on (road, trail, railway line, or forest) and predation sites where wolves had killed an ungulate. The tracking sessions were exported into the geographic information system (GIS) ArcInfo (Environmental Systems Research Institute, Redlands, California, USA) for data preparation and then were transferred to S-Plus (Insightful, Seattle, Washington, USA) for statistical analysis. We analyzed each pack separately and then pooled the data if the results were similar between packs.
Statistical analysisPath tortuosity. We calculated the tortuosity of wolf paths as log(L/R2), where L is path length and R is net displacement. We then used linear mixed-effects models (Pinheiro and Bates 2000) to determine how topography and human developments affected path tortuosity. We chose R2 as opposed to R because R2 commonly increases linearly with path length (Turchin 1998). We log-transformed the ratio of L/R2 to correct for heteroscedasticity and a right-skewed distribution of residuals across fitted values. Tortuosity may be scale dependent (Turchin 1998), so we measured tortuosity for path segments of three lengths: 0.5, 1.0, and 5.0 km. For each scale of analysis, we partitioned wolf paths into sequential segments of length (L). Path segments within each wolf path were probably correlated, so we included wolf path as a random effect in the mixed-effects model. Before partitioning the wolf paths into segments, we first minimized fine-scale wobble associated with GPS error by simplifying the wolf paths into a series of 100-m intervals.
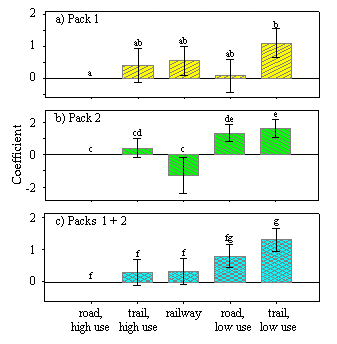
At each 100-m interval along the wolf paths, we collected attribute data from topographic and human development grids (pixel size of 25 m). Then, for each path segment of length (L), we calculated the median and variance of the attribute data collected within it. Attribute data included: aspect, elevation, distance to predation site, trail and road density, distance to high- and low-use roads and trails, distance to railway line, and distance to commercial accommodation. Topographic variables were derived from a digital elevation model (25-m resolution). Distance to predation site was defined as distance to a recent wolf-killed ungulate found along the wolf path (range of 0–13 km). Wolf paths without a predation site were assigned a distance of 20 km. Roads and trails were mapped with a hand-held GPS. All distances were measured in kilometers. Road and trail densities (km/km2) were measured as the total length of roads or trails within a circle (radius of 1 km) divided by the area of the circle. Commercial accommodations included both accommodations within and outside the town of Jasper. We created a preliminary global model and then used generalized additive models to identify nonlinear relationships between the explanatory and response variables (Venables and Ripley 1999). We log-transformed the median distance to predation site because all models with transformations had lower AICc values (Burnham and Anderson 1998) than did models without transformations. Consequently, as distance to kill site increased, its effect on path tortuosity decreased exponentially. At each scale of analysis, we identified the best linear regression model from a set of candidate models using AICc (Burnham and Anderson 1998). The magnitudes of regression coefficients depend on their scale of measurement. Therefore, to visually compare coefficients among models, we divided the regression coefficients by their standard errors such that all standard errors equaled 1.
Barriers to movement. We tested whether wolves circumvent and avoid crossing roads, trails, and a railway line by comparing the frequency with which wolf paths and a null model of simulated wolf paths crossed these features. For a valid comparison between observed and simulated paths, the shape of the simulated paths should be similar to the observed paths except for their responses to linear features, although other studies using this approach (e.g., Serrouya 1999, Dyer et al. 2001, Rondinini and Doncaster 2002) failed to test for similarity in shape of the simulated and observed paths. Accordingly, we created simulated paths based on the best of a series of random walk models. We will define four random walk models and then describe how we: (1) compared the models; (2) assessed how well the best model approximated wolf paths; (3) created a null model of simulated wolf paths using spatially explicit random paths paired with wolf paths; and (4) compared the frequency with which observed and simulated paths crossed linear features.
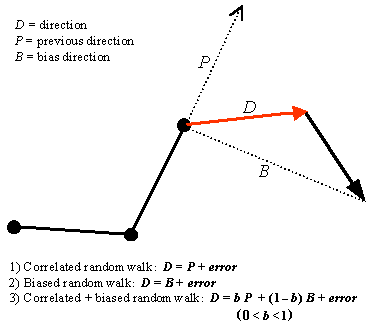
When testing and creating a random walk model, one first simplifies the movement paths into a series of points (Turchin 1998). Straight lines between successive points are called steps, and each step consists of a length and direction. The difference in direction of two steps creates a turn angle. The shape of a path depends on the magnitude and autocorrelation of these turn angles. Random walk models quantify the influence of directional persistence (correlated movement directions) and external (directional) bias on step direction. Thus, four common types of random walk models include uncorrelated, correlated, biased, and correlated + biased random walks (Turchin 1998, Schultz and Crone 2001). The simplest of these is the uncorrelated random walk, in which the predicted direction of each step is a random direction between 0 and 360 degrees. This model creates highly tortuous and circular paths and was not included in this analysis. We compared how well the remaining three random walk models predict wolf movement (Fig. 1). Correlated random walk models predict that the direction of the present step will equal that of the previous step and therefore will show stronger directional persistence and lower path tortuosity than the uncorrelated random walk. These models are common in the literature (e.g., Kareiva and Shigesada 1983) and are theoretically linked to models of population dispersal (Turchin 1998). Biased random walks model the influence of external factors on movement direction. In one type of biased random walk, organisms move toward a particular destination such as a nest, hive, den, or high-quality habitat. For this model, the predicted direction of travel is the direction toward the bias point (e.g., Schultz and Crone 2001). Finally, the correlated + biased random walk models the influence of both directional persistence and directional bias on an organism’s movement (Schultz and Crone 2001). This model predicts a weighted average of previous and bias directions. Weights (b and 1 - b) for this model were determined by iteratively adjusting the value of b from 0 to 1 until the maximum log-likelihood (minimum residual sum of squares) was attained. We defined step lengths and turning angles for the model comparisons by simplifying wolf paths into a series of points separated by 250 m and defined bias point as the end point of each path. We chose step lengths of 250 m to balance the strong negative autocorrelation inherent with shorter step lengths against the loss of resolution and the ability to detect the crossing of linear features with larger steps.
|
Fig. 1. Three types of random walk models based on directional persistence and directional bias.
|
To compare the relative fit of the three models to wolf paths, we calculated the difference between the observed and predicted directions of travel (residuals) for each wolf step, and summed these differences for all steps (residual sum of squares, RSS). Because directions are circular in nature (e.g., 370 degrees equals 10 degrees), we programmed the computer to adjust the residuals such that they lay between + 180 degrees (Jammalamadaka and SenGupta 2001). We identified the best of the three models using AICc. We assessed the overall fit of the best model by comparing the net displacement of random and wolf paths at lengths of 5, 10, and 15 km. To create random paths, we successively added steps to previous steps according to the equations defining each model. The direction of each step was determined by calculating the predicted direction of travel and adding a randomly selected residual (error). The error distribution of residuals for the correlated + biased random walk had a strong leptokurtic distribution with a mean and standard deviation of –0.02 + 34.53 degrees.
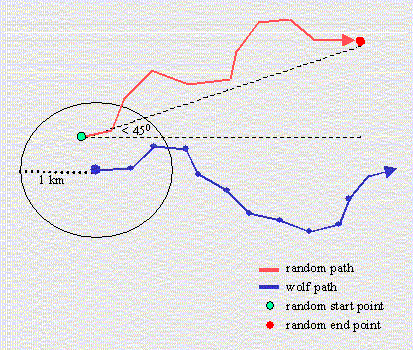
Using the best random walk model, we created 100 simulated wolf paths for each observed wolf path to define the expected number of times that wolves would cross linear features, given random movement. The simulated paths were paired with observed paths based on path length, start location, and end location (Fig. 2). The length of each simulated path was identical to that of its paired observed path. Start and end locations of the observed and simulated wolf paths were similar, but not identical, to avoid Type II error. The distribution of observed wolf paths was probably biased away from areas that the wolves avoided, such as areas with high road density (Mladenoff et al. 1995, 1999). We avoided this bias by selecting a random start location for the simulated wolf paths within 1 km of the observed start location. We then created an end location (bias direction) within 45 degrees of the observed bias direction. This angle represents the 75% confidence limits on the differences between the individual and average directions of observed wolf steps. We also avoided Type II error by creating start and end locations in areas where wolves would normally travel, such as lower elevations. For this, we compared the elevation, slope, and aspect of wolf and paired random locations using match case-control logistic regression (Hosmer and Lemeshow 2000). We then used this model to create a map in which each pixel contained a relative probability of wolf occurrence. When creating either a start or end location, we first generated two locations, determined the probability of wolf occurrence at each location, and then selected the location with the higher value. Consequently, the simulated paths were allowed to enter areas that wolves rarely traveled (e.g., elevations above 1500 m where only 3% of wolf locations occurred), but were more likely to occur in favorable habitat. As a final step in creating realistic simulated paths, we prohibited the paths from entering the town limits where wolves never travel. Once we generated 100 simulated paths for each observed path, we overlaid five types of linear features (high- and low-use roads and trails, and railway line) and counted the number of times that each observed and simulated path crossed these features. We chose 100 simulated paths per observed path as a balance between the intensive computational time required for spatially explicit simulated paths and precision gained by a large number of replicates. Imprecision from 100 simulated paths per observed path (as opposed to 1000 or 10,000 paths) was offset by the large number of observed paths (n = 177) and the resulting 17,700 simulated paths created for the study area.
|
Fig. 2. Each random path was paired with a wolf path in terms of path length. The start location was located within 1 km of the wolf start location and the end location lay within 45 degrees of the mean direction of the wolf path.
|
We determined whether wolves avoid crossing linear features by comparing the frequency with which observed and simulated wolf paths crossed high- and low-use roads, high- and low-use trails, and the railway line. The number of crossings per observed and simulated path fit neither a Gaussian nor a Poisson distribution because a high proportion of the paths did not cross linear features. Therefore, we changed the number of crossings per path to a binomial variable of either crossed or not crossed. We first tested whether wolves generally avoided crossing linear features by calculating the proportion of observed and simulated wolf paths that crossed each feature type and then running a t test (paired by feature type) on the differences between the observed and simulated proportions. Next, we tested whether wolves were more likely to cross some linear features than others. For this test, we used logistic regression and defined the response variable as whether or not the observed wolf path crossed a linear feature. The response (crossed = 1, not crossed = 0) was determined for each type of linear feature. Then, because the proximity of an observed path to linear features affected the likelihood of crossing, we also included availability as an explanatory variable. We maintained the pairing between the observed and simulated paths and defined availability as the proportion of the 100 simulated paths that crossed each type of linear feature. Thus, explanatory variables in the analysis were availability, feature type, and the interaction between availability and feature type. We conducted post hoc analyses on our final logistic regression model and tested for differences within feature type using Tukey’s multiple pairwise comparisons (α = 0.05; Bretz et al. 2001).
Over the course of two winters, we snow-tracked the two wolf packs over 1390 km (Fig. 3). We accumulated 91 tracking sessions for Pack 1 and 86 tracking sessions for Pack 2. The length of tracking sessions ranged from 0.5 km to 30 km, with a median length of 5.6 km. The wolves traveled on roads, trails, and railway lines 16% of the time and traveled through the forests, rivers, and meadows the other 84% of the time. Pack 1 traveled along the roads and trails (4 km on roads, 32 km on trails) less often than did Pack 2 (41 km on roads, 63 km on trails).
|
Fig. 3. Wolf paths through the study area from two winters of tracking (1999–2000, 2000–2001).
|
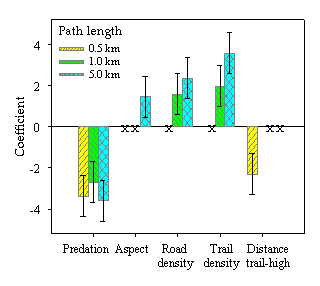
Path tortuosity. Models of path tortuosity were similar for both packs. Therefore, we pooled their data for analyses. At all three scales of analysis (path segments of 0.5, 1, and 5 km), the tortuosity of wolf paths increased near predation sites and either near high-use trails or within areas of high trail density (Fig. 4). Path tortuosity also increased in areas of higher road density at the larger two scales of analysis (1-km and 5-km path segments). Finally, path tortuosity increased with variation in aspect, but only for path segments analyzed at the 5-km scale.
|
Fig. 4. Standardized coefficients + one SE for linear mixed-effects models predicting path tortuosity. Explanatory variables are log(distance to predation site), variation in aspect, road density, trail density, and distance to high-use trail. Separate models were created for path segments of 0.5, 1, and 5 km. Explanatory variables with an X were omitted from the corresponding model.
|
At each of the three scales of analysis, the best model explained very little of the overall variation (adjusted r2 = 0.02, 0.02, and 0.27 for paths segments of 0.5, 1, and 5 km, respectively), but model performance increased for path segments of 5 km. The number of wolf paths (np) and number of path segments (ns) used in the analysis also varied greatly (np = 151, 126, 49; ns = 1149, 588, 77 for L = 0.5, 1, and 5 km, respectively). Although the predictability of the models was poor, all models explained more of the variation than did a model with a single intercept (ΔAICc = 15.1, 8.7, and 25.1 for L = 0.5, 1, and 5 km, respectively).
Barriers to movement. The correlated + biased random walk model more closely resembled wolf paths than did either the correlated or biased random walk models (Table 1). We assessed the validity of this model by comparing the net displacements of the wolf and random paths at 5, 10, and 15 km. The random paths were not significantly different from the wolf paths at 10 and 15 km, but had slightly smaller net displacements at 5 km (mean difference and 1 SE, 0.4 + 0.1 km). Large differences in net displacement between wolf and random paths of equal length might cause either Type I or Type II error. For example, if the start points originate far from human developments, the more tortuous random paths are not as likely to reach roads (Type II error); if start locations originate close to human developments, the more tortuous random paths are more likely to cross nearby roads (Type I error). A future improvement to null models of animal movement would be to create and compare separate random walk models from several categories of path lengths.
|
Table 1. Performance of three random walk models measured by differences between the observed and expected directions of travel for 2762 wolf steps. Wolf steps were 250 m in length. AICc values were calculated using the residual sum of squares (RSS) and the number of parameter estimates (K).
| ||||||||||||||||||||||||||||