|
|
|
Copyright © 2004 by the author(s). Published here under license by The Resilience Alliance.
Go to the pdf version of this article.
The following is the established format for referencing this article:
Moller, H., F. Berkes, P. O. Lyver, and M. Kislalioglu. 2004. Combining science and traditional ecological knowledge: monitoring populations for co-management. Ecology and Society 9(3): 2. [online] URL: http://www.ecologyandsociety.org/vol9/iss3/art2/
A version of this article in which text, figures, tables, and appendices are separate files may be found by following this link.
Synthesis, part of Special Feature on Traditional Knowledge in Social-Ecological Systems Combining Science and Traditional Ecological Knowledge: Monitoring Populations for Co-Management Henrik Moller1, Fikret Berkes2, Philip O'Brian Lyver1, and Mina Kislalioglu2
1University of Otago; 2University of Manitoba
- Abstract
- Introduction
- Monitoring Methods Used in Traditional Management Systems
- Potential Limitations of Traditional Population Monitoring
- Complementary Use of Science and Traditional Knowledge
- Discussion and Conclusions
- Responses to this Article
- Acknowledgments
- Literature Cited
Using a combination of traditional ecological knowledge and science to monitor populations can greatly assist co-management for sustainable customary wildlife harvests by indigenous peoples. Case studies from Canada and New Zealand emphasize that, although traditional monitoring methods may often be imprecise and qualitative, they are nevertheless valuable because they are based on observations over long time periods, incorporate large sample sizes, are inexpensive, invite the participation of harvesters as researchers, and sometimes incorporate subtle multivariate cross checks for environmental change. A few simple rules suggested by traditional knowledge may produce good management outcomes consistent with fuzzy logic thinking. Science can sometimes offer better tests of potential causes of population change by research on larger spatial scales, precise quantification, and evaluation of population change where no harvest occurs. However, science is expensive and may not always be trusted or welcomed by customary users of wildlife. Short scientific studies in which traditional monitoring methods are calibrated against population abundance could make it possible to mesh traditional ecological knowledge with scientific inferences of prey population dynamics. This paper analyzes the traditional monitoring techniques of catch per unit effort and body condition. Combining scientific and traditional monitoring methods can not only build partnership and community consensus, but also, and more importantly, allow indigenous wildlife users to critically evaluate scientific predictions on their own terms and test sustainability using their own forms of adaptive management.
KEY WORDS: adaptive management, catch per unit effort, community-based conservation, customary harvesting, indigenous people, population monitoring, sustainability, New Zealand, Canada.
Published: July 16, 2004
Traditional harvests of wild populations by indigenous peoples continue throughout the world (International Institute for Environment and Development 1994, IUCN 1997). These harvests are often needed for subsistence and may also be important social activities that help define the participants' cultural identity and provide links to their history, ancestors, land, art, and environmental philosophy (Kirikiri and Nugent 1995). Some of these customary uses of wildlife are controversial because they have been unsustainable, both historically (Martin and Klein 1984, Diamond 1992) and recently (Redford 1992). Mounting social, economic, and ecological pressures such as technology change, population growth, habitat destruction, species introductions, pollution, and climate change may exacerbate the impacts of customary harvests.
A common response of conservation agencies has been to advocate harvest prohibitions to safeguard biodiversity (New Zealand Conservation Authority 1997). However, in recent years some international conservation organizations have recognized that the imposition of harvest prohibitions is doomed to failure in the long term unless there is local support for conservation (International Institute for Environment and Development 1994, Posey 1996). Recent trends are toward participatory, inclusive, community-based approaches to conservation (Alcorn 1993, Hackel 1999, Berkes 2004). Recent reassertion of the rights of indigenous people has led to calls for the reinstatement of customary harvests in such places as northern Australia (Bomford and Caughley 1996), New Zealand (Moller 1996, New Zealand Conservation Authority 1997), Latin America (Alcorn 1993), and northern Canada (Treseder et al. 1999). However, it is not clear how these indigenous groups will manage their customary harvests, even if there is a willingness to conserve. Will they have the capacity or wish to use scientific approaches? Will they use their own traditional ecological knowledge and management systems? Alternatively, will they rely on a combination of the two, perhaps in the form of co-management?
In this paper, we argue that, instead of advocating prohibition, conservation efforts may be best directed to ensuring that customary uses are sustainable. Sustainability can be achieved through the complementary use by scientists of local and traditional ecological knowledge for joint management or co-management. Scientific research in support of sustainable customary use is therefore important and urgently needed.
Although the monitoring of resource population trends is a critical first component of the sustainable management of customary harvests of wildlife, it is not sufficient unto itself. Unsustainable use may persist if hunters choose not to respond to signals of declining resource levels or have no other options. However, reliable monitoring could signal potential overharvesting and then lead to an adjustment of hunting pressure to safeguard sustainability. Verification of sustainability predictions produced by mathematical models and adaptive management (Walters and Holling 1990) will also depend on the identification of a reliable population monitoring method. We argue that traditional ecological knowledge, which is based on learning by doing, is in many ways similar to adaptive management (Folke et al. 1998, Berkes et al. 2000). This paper therefore describes traditional ecological monitoring methods before considering whether they might be reliable enough to safeguard sustainability and promote learning by doing, on the assumption that hunters also have the will to adjust their harvesting based on the information they gather.
We ask how ecological science can play a complementary role in improving the environmental reconnaissance and sustainability of the customary use of resources. We do this by evaluating ways of combining science and traditional ecological knowledge to monitor populations. First, we review the monitoring approaches used in traditional ecological knowledge systems to make the point that indigenous harvesters are not simply optimal foragers, but may have ways of proactively responding to environmental signals. From this set of monitoring approaches, we analyze two in particular: catch per unit of effort and body condition. To generalize, we then consider examples of complementary uses of science and traditional ecological knowledge for population monitoring. This paper highlights case studies, mainly from New Zealand and northern Canada where we have firsthand experience. We define traditional ecological knowledge as "a cumulative body of knowledge, practice and belief, evolving by adaptive processes and handed down through generations by cultural transmission" (Berkes 1999:8). We use the term "co-management" to refer to a continuum of arrangements involving various degrees of power and responsibility-sharing between the government and the local community (Taiepa et al. 1997), and consider participatory research to be a form of co-management.
USED IN TRADITIONAL MANAGEMENT SYSTEMS
In scientific practice, ideal population monitoring methods are precise and easily measured in an objective and repeatable manner. However, conventional scientific research and monitoring can be expensive, often require specialized skills or technology, and are usually not practicable in the remote places in which customary resource use occurs. In such places, local communities possess neither a science capacity nor a tradition and trust in science. In addition, customary users are likely to be too busy to divert time to complicated monitoring methods, especially if the methods are independent of their harvesting.
Most traditional monitoring methods used by indigenous cultures are rapid, low-cost, and easily comprehensible assessments made by the harvesters themselves as they hunt. Hence, most of the known methods for monitoring populations are based on some aspect of the harvest or on observations related to harvests (Table 1). The harvest rate, or similar catch per unit of effort (CPUE) measurement, is the most practical population-monitoring index for customary resource users. We illustrate its practicality with two examples.
|
Table 1. Traditional methods to monitor populations.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
At the time of our study, the CPUE seemed to be the main mechanism for harvest decision making in the subsistence fisheries of the Cree people of James Bay in northern Canada. Cree fishers used 50-m gillnets of various mesh sizes and kept a mental note of their harvests and their CPUE relative to expected CPUE based on previous experience; decisions about when and where to move were based on this collected information. The CPUE was the primary mechanism driving three practices: concentration of effort, pulse fishing, and the use of a mix of gillnet mesh sizes. The CPUE shaped decisions regarding which nets to use, how long to keep fishing in a given spot, when to relocate, which species to target, and how often to visit distant but high-CPUE areas. The Cree fishers monitored other environmental signals as well, including the species composition of the catch; size distribution; body condition, which was considered important as an indicator of health; and reproductive condition (Berkes 1999).
The Rakiura Maori, New Zealand's southernmost group of indigenous people, travel to 35 islands each autumn to harvest the chicks of the sooty shearwater (Puffinus griseus). This "muttonbirding" is important for the cultural identity of the group as well as for the economic return gained from the sale of the chicks. Harvesters primarily monitor the long-term well-being of the birds, which they call titi, from the rate at which they can catch chicks (Lyver et al. 1999, Kitson 2004). Declining catch rates in the absence of a major change in body condition (Lyver 2002), harvest intensity, or breeding habitat in New Zealand led customary users to conclude the existence of distant influences, probably during migration to or from the north Pacific. Many titi harvesters keep written records of catch totals and weather or moon conditions during each hunt, in many instances going back for decades, e.g., to 1927 in one case. Most titi harvesters base their assessment of trends in abundance on whether they can still get their target daily total catch or "tally," whether that total is achieved in about the same time, or whether it is gathered from about the same area of breeding ground, sometimes measured indirectly as how far birders must go from their base to complete the harvest. Harvest rate guides short-term decision making to optimize returns. Anticipated harvest rate based on "a quick look-see" is a key cue for deciding whether to mount a prolonged harvest expedition on a given night (the "starting rule") and when to discontinue harvest as moonlight triggers the return of the chicks to their burrows (the "stopping rule").
A second method by which some traditional management systems monitor populations is the use of body condition. It has been known for some time that indigenous hunters of caribou (Rangifer tarandus) who belong to a number of distinct groups from Labrador to Alaska have systems of monitoring caribou fat content (Berkes 1999:108, Kofinas et al. 2004). Some of these assessments are based on the size and gait of the live animal before it is selected for harvesting; other are made during butchering as a retrospective assessment of herd condition. Cree and Dëne hunters and fishers regularly check the fat content of the animals they harvest. They pinch the undersides of the bodies of geese and ducks, assess the mesentery fat of fish when cleaning them, and check the fat content of large mammal species.
Three indicators for monitoring of body condition, i.e., back fat, stomach fat, and marrow color, top the list of some nine indicators of a healthy caribou used by Porcupine Caribou hunters (Kofinas et al. 2002). Monitoring caribou fat makes sense as an index of the health of the individual animal. It could also make sense as an index that integrates a number of factors at the ecosystem level, such as the condition of the feeding range and population dynamics. In fact, some Cree hunters were using caribou body fat as a way of tracking range extension into the James Bay area in the 1980s. They considered the observation of fat (and healthy) caribou as indicating the use of new feeding grounds, but did not seem to interpret body condition in terms of population size (F. Berkes, personal observation). Kofinas et al. (2002) report that hunters of the Porcupine Caribou herd considered poor body condition an indication of overpopulation. However, hunters and elders from the Chipewyan Dëne community of Lútsël K'é in Canada's Northwest Territories do not link fat on barren-ground caribou (Rangifer tarandus groenlandicus) to population abundance, but rather use it as an indicator of population well-being and range conditions (P. Lyver, personal observation). Lútsël K'é hunters use the caribou bulls harvested during the fall and the cows harvested during the winter and spring to assess temporal and spatial variations in body condition (P. Lyver and Lútsël K'é Dëne First Nation, unpublished manuscript).
Table 1 summarizes a number of other methods used by traditional hunters to monitor population abundance. Observation of breeding success is one of them. In late summer, before nesting populations of Canada geese (Branta canadensis) start their migration south, Cree hunters in James Bay monitor breeding success for the year by noting the number of young per nest. Because there are local prohibitions against disturbing nesting geese, this monitoring takes place only at the edges of nesting areas as hunters pass by on the way to other activities. By the time the geese are ready to migrate in September, most hunters have shared this monitoring information and can predict a good or a poor hunt. When the harvesting starts, hunters also start monitoring the average young per adult couple and thus the likely productivity of the hunt. All this helps them plan how much time and effort to put into the goose hunt, as compared to other options such as fishing or big game hunting (Berkes 1982). Shifting effort to other resources in this manner may help reduce the pressure on the resource population that is experiencing a low.
Hunters also form impressions of population status through their various senses. "Eyeballing" the numbers of birds, game, and even fish, e.g., salmon in upstream migration, is common (Swezey and Heizer 1977), and a number of groups of indigenous hunters assess population numbers by monitoring the density of tracks. Maori sometimes refer to sensing their environment through "touch, feel, and sight" (H. Moller and S. Sunseri, unpublished manuscript). Other senses are also used. Titi harvesters have been known to assess the presence or absence of birds by smell and sound (Heaslip 2002); Cree hunters sometimes judge the numbers of the Canada goose population in staging areas by the amount of noise they make (F. Berkes, personal observation). The results may be more reliable if such assessments are based on a community consensus rather than predominantly individual endeavors. For example, some tribal groups in India engage in large-scale animal hunts (Gadgil et al. 1993).
One of the characteristics of traditional monitoring is that observers tend to note unusual rather than average patterns and occurrences. For example, community-based observations of climate change in various parts of the Arctic say little about averages but indicate a consensus that weather is more variable and less predictable, with an increased frequency of extreme weather events (Krupnik and Jolly 2002). Indigenous observers similarly note unusual patterns in animal distributions, strange behavior, diseased animals, and breeding failures. For example, massive titi food failure events known as "kiaka years" are long remembered and recounted by Rakiura Maori titi harvesters (Hunter et al. 2000). Changes in the frequency of such unusual events are often interpreted as signs of long-term alterations in ecosystems or resource levels.
Observations of species mixes or assemblages may also provide important information on upcoming population change. The cabbage tree or ti (Cordyline spp.) on the North Island of New Zealand started dying for an unknown reason in the mid-1980s. As the decline worked its way south, the New Zealand Department of Conservation commissioned a report on Maori knowledge of the condition. The report noted the observation that ti "need to be part of a vegetation complex, rather than all alone in a paddock" (J. Williams and T. Chrisp, unpublished manuscript). Some months later, scientists isolated the passion vine hopper as the disease-carrying vector. The hopper does not fly more than 1.5 m above ground and is inhibited by surrounding vegetation that also protects ti against accidental damage to the lower trunk. The response of ti to injury is to grow another head at the injury site, and it is only at the base of a leafy growth that the hopper can penetrate the cortex and exchange fluids (P. Simpson, personal communication).
Table 1 presents the monitoring methods as though they were independent of one another. However, customary users often combine different methods or integrate them for cross-checking purposes and better understanding. Titi harvesters adjust their behavior by prolonging hunting or by hunting in new areas or at suboptimal times to buffer harvest returns against declining CPUE. However, many are still aware of a declining resource. This awareness is informed by visually assessing the number of adults returning at night, the smell and sound of the birds, and the lower total catch rates. The Rakiura Maori harvesters we interviewed were aware that a previous ecological pattern had changed in the last decade. Formerly, years of high and low chick abundance used to be correlated with fat and thin chick years, respectively. However, the recent occurrence of seasons with low numbers of fat chicks signaled an environmental perturbation to the system that affected the number of adults in the population rather than their ability to feed chicks (Lyver 2002). This is the traditional ecological knowledge equivalent to simple multivariate analysis.
Although none of the traditional methods in Table 1 are quantitative, they do provide a set of "rules of thumb" (Gadgil et al. 1993) for monitoring populations. This brings us to the question of the extent to which these rules of thumb provide a satisfactory guide for assessing populations and for sustainable management. Before addressing this question, it is necessary to examine in detail some limitations of traditional monitoring methods, using catch per unit of effort and body condition indices as examples. It should be noted that aspects of the following analysis are also applicable to scientific monitoring.
TRADITIONAL POPULATION MONITORING
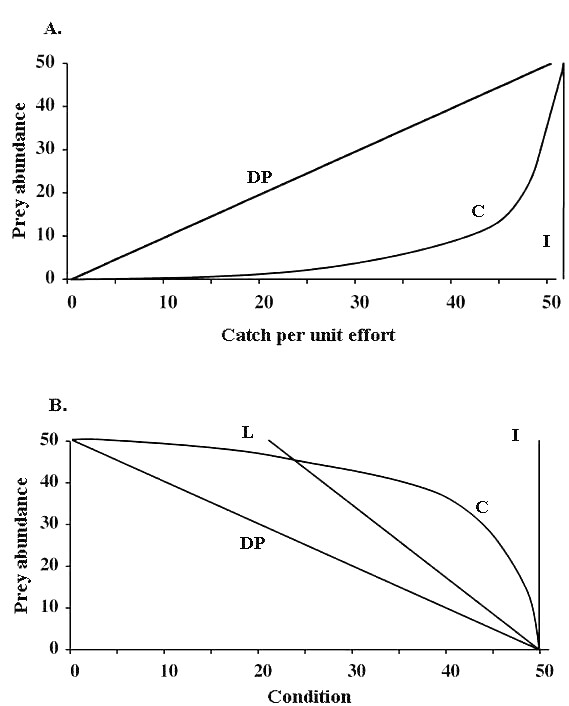
Despite the widespread use of harvest rate indices to guide wildlife management, there have been few demonstrations of their precision as relative population indices or of calibration of catch per unit effort (CPUE) against actual prey density. First principles suggest that the null model ("I" in Fig. 1A) of complete independence of CPUE from prey density will indeed be rare, but the usual assumption that harvest rate is linearly related and directly proportional to density (line "DP" in Fig. 1A) is probably far too simplistic in most circumstances. It assumes that a 50% reduction in harvest rate indicates a 50% reduction in abundance. This is akin to assuming random encounter rates between hunter and prey, so that a doubling of prey numbers will double the encounter rate and consequently double the CPUE. In practice, harvest rate is more likely to be curvilinear in relation to prey density, usually because CPUE levels off at high prey density (line "C" in Fig. 1A). Curvilinearity will arise partly because of constant handling and constraints on travel time, or because the hunter becomes "saturated" when confronted with a hyperabundance of prey. Their rate of capture is thereby set at the physical maximum constrained by the mechanics of killing and/or processing prey rather than by the time required to find and kill the next prey.
|
Fig. 1. A notional calibration of (A) catch per unit effort (CPUE) and (B) body condition against prey population abundance. The abundance scale is the proportion of the population carrying capacity, and the CPUE and body condition measure is the proportion of the maximum value observed. "I" represents density independence, "DP" shows a direct proportionality between the index and abundance, and "C" shows a curvilinear relationship. Line "L" in Fig. 1B depicts a linear relationship between the index and abundance, but does not directly represent proportionality because body condition does not fall to zero at carrying capacity. The relationships in Fig. 1B could apply equally to breeding success when used as a factor for monitoring.
|
For instance, "handling time," which includes retrieving, butchering, and processing the food, and "travel time" to the location of the hunt and back to base with the produce will usually be relatively independent of prey density. If handling and travel times are relatively long compared to the cumulative time encountering prey, a large variation in prey density will have relatively little impact on overall CPUE. A hunter pursuing aggregated prey is saturated and will kill about the same number of prey irrespective of the herd or flock size. Saturation of fishing gear, where all hooks are taken or a net loses efficiency as it fills up, may produce a similar curvilinear relationship in CPUE calculations based on fish per line or per net per day. In the case of the Rakiura Maori, a harvester encountering a cluster of emerged titi chicks can catch only three or four of them before the others escape into nearby breeding burrows. Similar disturbance effects are likely to occur in other systems.
Saturation of a hunter's efficiency leads to the CPUE becoming nearly independent of prey density when prey density is high, as depicted at the upper right of line "C" in Fig 1A. However, harvesters may adjust their own behavior at very low prey abundance in ways that make the overall mathematical relationship between harvest rate and abundance nonlinear at the lower left end of line "C" in Fig 1A. In particular, harvesters use their traditional ecological knowledge to target particular high-density places or "hot spots" of prey. Rakiura birders not only target particular areas on their island where in the past catches have been good, i.e., greater chick density, but they also track temporal variations in the location of high-density areas. During the second part of the harvesting season, birders maximize their CPUE by hunting during optimal weather, i.e., rain, wind, and low light, and they also vary the location of the hunt according to wind direction (Lyver 2002). These adjustments "cream off" high-density places and thereby trigger a rapid increase in the harvest rate even at very low overall prey abundance (line "C" in Fig 1A).
Harvesting only at high-density places or aggregations of prey raises a generic concern that the use of CPUE to monitor changes in overall population abundance may not reflect increasing or decreasing prey distribution. If the harvest rate is nearly always measured at high-density places or in the center of the population's distribution, the harvester may be unaware of changes in the overall distribution of the prey population or in the ratio of high-density places to low-density places within that overall range.
A similar nonrandom sampling problem results from temporal variations in resource availability and consequent adjustment by harvesters of when they will pursue their prey. For example, Rakiura birders will not even start a hunt if conditions suggest that a low harvest rate will ensue. Furthermore, they will suspend the hunt if conditions deteriorate, e.g., rain or wind ceases, the moon rises, during the hunt (Lyver 2002). In addition, most hunters stop once they have caught a target total of chicks for the night. Because this tally is most likely obtained in good hunting conditions when relatively high numbers of chicks have emerged from their breeding burrows, the sample of harvest rates recorded will be biased toward the short time periods of high prey availability. This creates practical difficulties when using harvest rate to monitor low populations and, more subtly, a bias from the incorporation of nonrandom sampling times. The effect of temporal and spatial nonrandom sampling by efficient hunters is to buffer changes in CPUE indices from changes in prey abundance. The more skillful the hunter, the poorer the CPUE index will be as a monitor of population abundance.
Monitoring populations based on body condition or productivity not only presents similar potential pitfalls of nonrandom sampling in place and time, but also introduces the added complication of the selectivity of harvesters who target the fatter animals, which leads to nonrepresentative sampling. Caribou harvesters target the fattest animals because they are nutritionally more valuable and considered healthier (Kofinas et al. 2004). Similarly, titi harvesters are skilled at finding and selecting the fattest chicks (Hunter et al. 2000). Historically, the fat itself was highly sought after for its nutritional value and to facilitate the preservation and transport of harvested birds; fat birds are still more marketable and considered to taste better. The degree of selectivity of titi chicks varies with the harvester and the number of chicks available at a given season or given time of year. Therefore, a simple measure of the fat or body condition of harvested birds may not vary proportionally with the absolute change in body condition within the population.
Even if a reliable relative index of body condition can be obtained from a harvester's sample, the interpretation of the meaning of changes in body condition is potentially problematic because of density dependence. Furthermore, even if the condition of individuals linearly relates to density, it is most unlikely to index directly proportional changes in abundance (line "DP" in Fig 1B). A linear relationship (line "L" in Fig 1B) may occur, but a curvilinear one ("C" in Fig 1B) is more likely. Both lead to increased body condition when the population is falling, presumably because of the release from density-dependent pressure on food supplies or disturbance competition. Increasing body condition could therefore signal an unsustainable overharvest, and falling body condition could signal overpopulation and potential range damage by overgrazing.
Increased body condition is frequently interpreted as sign of healthy individuals (Kofinas et al. 2004), but this is not the same as saying that the population is healthy and either stationary or increasing. High body condition can be an indicator of population well-being only if body condition is independent of density ("I" in Fig 1B), and if body condition affects vital rates in the population such as reproduction, survival, emigration, or immigration. Independence from density suggests that factors other than food supplies, e.g., competition for breeding space, limit the population.
There is a close relationship between the harvesters' own qualitative assessments of caribou body condition and quantitative measures by researchers, and both types of index predict pregnancy rates (Lyver and Gunn 2004). However, there is no certain evidence of the density independence of caribou body condition. Researchers have found that the size of the George River caribou herd primarily regulates the amount of forage available on the summer range (Manseau et al. 1996) and inferred that this could be responsible for the declines in body condition when the herd nears carrying capacity. Caribou fat index may be more useful as a short-term index of fluctuating environmental conditions for a given season, or at least within the time span of the "physiological memory" of the animals being hunted. It may have relatively little value for long-term population monitoring per se. Just as in science, traditional monitoring methods must not rely on the measurement of a single variable without integration of knowledge of the overall ecological system.
Even where body condition can be demonstrated to affect a population parameter, population regulation and compensatory adjustments in other population parameters could weaken or completely counteract such changes. This compensation would weaken the ability of body condition to predict future population changes. For example, changes in titi chick body condition undoubtedly affect recruitment (Hunter et al. 2000), but compensatory changes in older birds potentially compensate for changes in the parameters affecting chick and young stages (Moller 2004).
Monitoring the breeding success of populations (Table 1) has many of the same potential complications as the body condition index. Density dependence in breeding success of the types depicted in Fig. 1B is probable, and variations in breeding success can only indicate longer-term consequences at the population level if other compensatory changes do not mitigate the effects of increased or decreased production. Higher breeding success can therefore give little indication, or even a very misleading signal, of where the population is heading.
SCIENCE AND TRADITIONAL KNOWLEDGE
The preceding two sections emphasized both the potential value and the pitfalls of traditional monitoring methods. We turn now to a consideration of how ecological science and traditional ecological knowledge could play complementary roles to improve the prospects for sustainable customary harvests.
As seen in Fig. 1, scientific study to reveal the shape of the calibration between prey abundance and catch per unit effort (CPUE), body condition, or productivity can immediately assist research partnerships by determining whether a change in traditional monitoring index would be expected with a change in actual density. For example, there is no prospect whatever of Rakiura Maori harvesters using the slow, specialized, and expensive "burrowscope," a miniature video camera on the end of a flexible tube inserted down into the burrow (Lyver et al. 1998) by the titi research team to count the number of chicks. Indeed, the birders were often bemused by and skeptical of the burrowscope and its operators after watching our laborious fieldwork. The burrowscope did indeed miss several chicks (Hamilton 2000), and thorough inspection of burrows is several times slower than the birders' own method of finding chicks using a wire. Our response has been to put considerable effort into calibrating the traditional methods against actual abundance measures established using the burrowscope so that the advantages and potential limitations of the former are exposed (Kitson 2004). Research co-management agreements often secure data-sharing agreements, but divergent inferences from each other's knowledge system, i.e., science vs. ethnoscience, could be resolved by intensive calibration studies.
An unexpected bonus of such studies for titi CPUE has been increased consensus among birders harvesting in different family territories about what is happening to the resource. Families harvesting an area with abundant birds per harvester are operating at the upper end of curve "C" in Fig. 1A, and so have noticed little change in the harvest rate over the past decades of declining population. Others have been noticing declines in CPUE because they have relatively few birds per harvester at the lower end of "C." Thanks to calibration, the claim that the bird population was declining was more widely accepted, and this may now lead to a more cohesive community evaluation of whether or not to manage the situation.
The Rakiura Maori asked their science team to devise a practical but statistically robust monitoring protocol that could be applied by the birders themselves. Computer simulations using measures of variability in harvest rates between individuals, areas, and times will gain statistical power from the predictions of a panel of traditional experts including various numbers of birders who report their harvest rates for the community's assessment of population trends. Research has shown that the harvest rates of novices and very old birders are relatively lower than those of experienced middle-aged groups, so it is necessary to adjust the makeup of the panel or make some statistical accommodation for these different rates.
Retaining traditional monitoring methods is potentially crucial because they provide one of the few channels through which customary users can scrutinize science. If scientific methods cannot be tested, they are less likely to be trusted, and their findings will probably not be acted upon. The common perception among customary users that scientists are arrogant comes from the presumption that science provides the "gold standard" against which traditional knowledge system must be judged. However, a partnership of equals requires reciprocity and humility (Berkes 1999), and integrating traditional monitoring methods may serve to ameliorate existing conflicts. We will discuss two cases from northern Canada to further our argument that ecological science and traditional monitoring may play complementary roles.
The James Bay and Northern Quebec Agreement of 1975 devolved management authority over beaver (Castor canadensis) to Cree hunters and their organizations. Since at least the 17th century, the Cree have managed beaver populations through stewards who are senior hunters and family heads (Francis and Morantz 1983). Typically, these stewards make decisions about how many beaver to harvest per year in the territories under their control. Under the agreement, this right was formalized, and the stewards were given access to aerial survey data by the provincial resource management agency. By the mid-1980s, Cree hunters were avidly using these data. Aerial surveys provided an accurate count of the number of beaver lodges over a large area, but could not determine if beaver actually occupied a given lodge. The stewards, in turn, knew what proportion of the lodges in a given area were actually occupied, but did not have good data on the total number of lodges in their territories, which are often several hundred square kilometers in size (Berkes et al. 1989; F. Berkes, personal observation). The combination of scientific monitoring data and hunters' local observations provided optimal monitoring information to manage the resource.
The second case from northern Canada concerns the Beverly-Qamanirjuaq Caribou Management Board, a co-management body set up in response to the perceived sharp declines in the 1970s in the caribou herd of the same name. According to aerial surveys over a large territory, the herd was judged to have declined to dangerously low levels, and a co-management body was set up to bring together government managers and caribou hunters belonging to three distinct indigenous groups: the Inuit, the Dëne, and the Cree. However, what started as a body for the "education" of indigenous hunters who may have been responsible for overhunting turned into a highly successful and long-enduring cross-cultural forum for joint problem solving (Kendrick 2000). Shortly after two-way communication started among the indigenous hunters and government managers, it was discovered that the caribou herd had split and that part of the herd was outside the area being surveyed. Once the "caribou crisis" was over, the co-management group could focus its efforts on discussing allocations and on setting up joint research projects and population monitoring programs that could satisfy the requirements of both the users and the government managers (Klein et al. 1999). However, progress has been slow, and some conflict remains. There has been much debate on the cultural acceptability of radio-collaring the caribou used to monitor movements and to guide biologists to the herds for the measurement of cow:calf ratios.
All our cases show that, when it comes to determining the nature of the problem, there is a surprising level of agreement between traditional ecological knowledge and science (Lyver 2002, Newman and Moller 2004), particularly in the area of primary observational evidence of changing patterns in the environment. However, because of their different world views and other factors, scientists and local communities often disagree about what is causing the problem and how to go about solving it. Clearly, scientific and indigenous knowledge differ in their approach to population monitoring, and these differences are the source of many disagreements. However, an alternative view of these differences might be to treat them as different areas of expertise that complement rather than contradict each other.
There is a bicultural component to the conservation strategy for cabbage trees in New Zealand because both Maori and Pakeha, i.e., nonindigenous New Zealanders, bring different world views to the subject while sharing the same goal. The combined approaches of Western science and of indigenous knowledge or matauranga Maori offer a far stronger prospect than either approach on its own (Simpson 2000:299).
In such a partnership, science can supply data from large areas, as seen in the caribou, beaver, and titi examples. Science can establish more spatial generality, provided that sampling is designed at the appropriate scale and well replicated. Data generated by science are quantitative and, within limits, objective. Scientific monitoring methods offer the advantage of decoupling the sampling from the harvest itself. An index based on harvest rates cannot be used in nontreatment or unharvested areas. Because most, although not all, of the monitoring measures in Table 1 are related to harvesting and processing, they cannot track resource abundance where the harvest does not take place. From the point of view of formal science, this makes it impossible to use an experimental or adaptive management approach to isolate harvest effects from other potential ecological impacts.
Science can serve a collaborative partnership by providing a study of causation. Scientists think in terms of cause-effect relationships determined entirely by biophysical mechanisms, whereas those who base their actions on indigenous knowledge sometimes attribute events or environmental changes to very different mechanisms. The latter often use metaphorical language when discussing the reduction or disappearance of populations, and their discourse on population phenomena is therefore sometimes strikingly different from scientists. For example, Kofinas (2002) documents a case in which the failure of caribou to overwinter nearby was considered a result of the recent death of a hunter who had had special powers of communicating with the caribou. Such explanations could never be evaluated on scientific grounds. However, science complements traditional ecological knowledge by testing causation, as illustrated by the titi example.
The transequatorial migration of titi adults increases their chance of encountering fishing vessels in the mid and north Pacific, making them vulnerable to accidental drowning in nets and also to global climate perturbations that affect wind and food. Such impacts could never have been investigated by traditional ecological knowledge alone, because this knowledge is based almost entirely on chick numbers and conditions at the breeding colonies in the southern hemisphere (Lyver et al. 1999). International cooperation using state-of-the-art scientific technology is under way to track adults and correlate survival with bycatch and climate change to identify the mechanisms of decline. However, the scientific search for the causes of decline might not have happened if the birders had not become concerned (Davis 2001, Moller 2001). Traditional ecological knowledge also identified the most probable proximate cause for declines and thereby focused the scientific investigation. Meticulous records covering more than two decades indicated that adult numbers had declined probably because of climate change rather than food shortages that had restricted breeding and chick growth (Lyver 2002). The contributions of traditional ecological knowledge to the partnership included the ability to help detect such important changes and pave the way for the formulation of useful scientific hypotheses (Newman and Moller 2004).
The diachronic nature of traditional ecological knowledge provides robust temporal perspectives and baseline information that is on a local scale and based on a particular place. Such information is often crucial for the conservation of long-lived species with low population turnover. A review of perturbation experiments in ecology showed that most of the experiments provided very weak inference because they usually run for less than a single generation of the response species (Raffaelli and Moller 2000). Traditional ecological knowledge can stretch the time scale to living memory and, in some cases, oral history. However, traditional harvesters may be prevented by social constraints from observing neighboring territories, or may not bother to do so simply because they are harvesting effectively in their own location. Indeed, familiarity with one's own place is itself a key determinant of harvest success and therefore a disincentive to sample elsewhere. Good science, on the other hand, gains power by sampling at other places or at nonharvest times of the life cycle to investigate harvesting and other population impacts.
The inclusion of traditional ecological knowledge has the potential to greatly increase the sample size. For example, annual harvest estimates from the George River caribou herd are approximately 40,000 animals; for the Bathurst herd, approximately 18,000; for the Porcupine herd, 2600–3900; and for the Western Arctic herd, 15,000–20,000 (Porcupine Caribou Management Board 2001; Kofinas et al. 2004; J. Dau, personal communication). The large number of samples that results may improve the accuracy of measurements, especially where there is a close relationship between the traditional body fat index scores and well-quantified scientific ones (Lyver and Gunn 2004). In contrast, the Yukon territorial government has conducted a detailed and quantitatively based analysis of the body condition of just 15–20 cows biannually for the past 10 yr and extrapolated the condition of the entire herd from the findings (Kofinas et al. 2002).
Another area in which traditional ecological knowledge can serve the partnership concerns the observation of extreme events and long-term adaptive strategies based on these observations (Riedlinger and Berkes 2001, Krupnik and Jolly 2002). Because of its diachronic nature, traditional ecological knowledge is less likely to miss occasional extreme events, whereas science may miss the event altogether because of a short sampling duration. On the other hand, a shift or trend in the average pattern may be ecologically more important or signal different concerns than a shift in extreme events. Consequently, using science to focus on shifting averages can complement the understanding of shifts in extremes or in environmental predictability gained from traditional knowledge.
Still another area concerns the "knowledge-practice-belief" complex embodied in traditional ecological knowledge (Berkes 1999). Deliberate inclusion of people and relationships in traditional ecological knowledge may provide a dimension missing from conventional science, thus humanizing ecology (Berkes 1999), and providing what the Maori would call "science with a heart."
Table 2 summarizes five areas of complementarity arising from the above discussion. These five points summarize the areas in which science and traditional ecological knowledge may strengthen one another with respect to population monitoring, and some of these points may be applicable to other areas of environmental science as well (Colding and Folke 2001, Riedlinger and Berkes 2001).
|
Table 2. Areas of complementarity between science and traditional ecological knowledge for population monitoring.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
A more holistic approach to population monitoring may consider taking into account factors such as those given in Table 2. On the one hand, traditional ecological knowledge alone cannot address all of the problems of population monitoring. On the other, problems of ecological scale and consequent difficulties in sampling and experimentation limit the predictive power of science (Raffaelli and Moller 2000). There is a growing recognition that conventional scientific approaches may be insufficient in the face of complexity. Problems of complex adaptive systems involving human uses and impacts often cannot be separated from issues of value, equity, and social justice; they require participatory approaches in which scientists need to work with local people (Ludwig 2001).
The reduction of complexity to a few simple rules (Gadgil et al. 1993) can produce remarkably robust outcomes. In some cases, rules of thumb may be more important than numerical precision. Traditional monitoring methods such as those listed in Table 1 are often used in combination with one another. "See, touch, feel" monitoring may not be considered "good science," but in combination with other ways of sensing populations, and backed up by a modest amount of good monitoring science, it has the potential to help the search for sustainability. Seemingly complicated problems can sometimes be solved by a few simple, often intuitive, decision rules, as demonstrated by fuzzy logic (Mackinson 2001).
In conclusion, some combination of science and traditional ecological knowledge is probably needed to monitor customary harvests. Some cases of customary resource use and management are based on indigenous knowledge and management practice (Colding and Folke 2001). These traditional systems, founded on a local understanding of ecosystems and developed by trial and error over time, have certain similarities to adaptive management (Berkes et al. 2000). However, they have not always been effective in protecting resources, which is also the case with scientific management (Gunderson and Holling 2002). The emergence of adaptive management, or "learning by doing" (Walters and Holling 1990), as a method of understanding ecosystems may be considered an indirect acknowledgement of the similarities between traditional management and scientific management.
All of our case studies emphasize the importance of gaining reliable information at temporal and spatial scales appropriate for monitoring natural resources (Folke et al. 1998). Traditional knowledge and science can complement each other by providing information about different parts of these spatial and temporal continua. Traditional knowledge often provides impressive temporal depth, but could be misled by focusing on the richest prey patches. Science can inform about key changes outside harvested areas that affect the resource and is potentially better able to determine why changes occur and thereby encourage management intervention to alter outcomes.
Traditional and scientific management systems may prove complementary, and differences between them more theoretical than real. Because science is expensive and many customary harvests take place in developing countries or in poor regions, simple, rapid, and inexpensive yet robust monitoring methods are needed. There are many practical difficulties in mounting the actual research and building trust between the researchers and the traditional harvesters they wish to support (Moller 2001). Involving the harvesters themselves by using their own monitoring methods or inviting their participation and partnership with scientific monitoring is much more likely to lead to the application of the results and altered harvest practice where needed for sustainability. Scientific methods must be congruent with traditional management and acceptable to local resource users. Complementing the limited scientific monitoring information in such regions with traditional ecological knowledge is potentially workable and cost-effective (Johannes 1998), and adaptive management may provide a framework for joint use.
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a comment, follow this link. To read comments already accepted, follow this link.
PL's and HM's research was funded primarily by the New Zealand Foundation for Research, Science and Technology to the Rakiura Maori and to PL for post-doctoral study in Canada. FB's research was supported by the Social Sciences and Humanities Research Council of Canada (SSHRC), and MK's by the Canada Research Chairs program (http://www.chairs.gc.ca). The collaboration for this paper was made possible by the University of Otago, William Evans Fellowship to FB. We thank Sicily Sunseri for editing and help with the literature. Allyn Stearman and an anonymous reviewer made helpful suggestions on an earlier draft of the manuscript.
Alcorn, J. B. 1993. Indigenous peoples and conservation. Conservation Biology 7:424-426.
Berkes, F. 1982. Waterfowl management and northern native peoples with reference to Cree hunters of James Bay. Musk-Ox 30:23-35.
Berkes, F. 1999. Sacred ecology, traditional ecological knowledge and resource management. Taylor and Francis, Philadelphia, Pennsylvania, USA.
Berkes, F. 2004. Rethinking community-based conservation. Conservation Biology 18, in press.
Berkes, F., J. Colding, and C. Folke. 2000. Rediscovery of traditional ecological knowledge as adaptive management. Ecological Applications 10:1251-1262.
Berkes, F., D. Feeny, B. J. McCay, and J. M. Acheson. 1989. The benefits of the commons. Nature 340:91-93.
Bomford, M., and J. Caughley, editors. 1996. The sustainable use of wildlife by Aboriginal peoples and Torres Strait islanders. Australian Government Publishing Service, Canberra, Australia.
Colding, J., and C. Folke. 2001. Social taboos: "invisible" systems of local resource management and biological conservation. Ecological Applications 11:584-600.
Davis, J. 2001. Te kaha te hikoi o te Tangata. In M. Howard and H. Moller, editors. He Minenga Whakatü Hua o Te Ao [Sustaining the fruits of the land]: proceedings of a hui. (Murihiku Marae, 2000). Available online at: http://www.otago.ac.nz/Zoology/hui/Main/Talks2/Davis.htm.
Diamond, J. M. 1992. The rise and fall of the third chimpanzee. Vintage Books, London, UK.
Folke, C., F. Berkes, and J. Colding. 1998. Ecological practices and social mechanisms for building resilience for sustainability. Pages 414-436 in F. Berkes and C. Folke, editors. Linking social and ecological systems. Cambridge University Press, Cambridge, UK.
Francis, D., and T. Morantz. 1983. Partners in furs: a history of the fur trade in eastern James Bay, 1600-1870. McGill-Queen's University Press, Montreal, Canada.
Gadgil, M., F. Berkes, and C. Folke. 1993. Indigenous knowledge for biodiversity conservation. Ambio 22:151-156.
Gunderson, L. H., and C. S. Holling, editors. 2002. Panarchy: understanding transformations in human and natural systems. Island Press, Washington D.C., USA.
Hackel, J. D. 1999. Community conservation and the future of Africa's wildlife. Conservation Biology 13:726-734.
Hamilton, S. A. 2000. How precise and accurate are data obtained using an infra-red scope for burrow-nesting sooty shearwaters (Puffinus griseus)? Journal of Marine Ornithology 28:1-6.
Heaslip, T. 2002. Impressions of the Muttonbird Island 2002. Titi Times 11:15.
Hunter, C. M., H. Moller, and J. Kitson. 2000. Muttonbirder selectivity of sooty shearwater (Titi) chicks harvested in New Zealand. New Zealand Journal of Zoology 27:395-414.
International Institute for Environment and Development. 1994. Whose Eden? An overview of community approaches to wildlife management. IIED, London, UK.
IUCN Intercommission Task Force on Indigenous People. 1997. Indigenous people and sustainability. IUCN, Gland, Switzerland.
Johannes, R. E. 1998. The case for data-less marine resource management: examples from tropical nearshore fisheries. Trends in Ecology and Evolution 13:243-246.
Kendrick, A. 2000. Community perceptions of the Beverly-Qamanirjuaq Caribou Management Board. Canadian Journal of Native Studies 20:1-33.
Kirikiri, R., and G. Nugent 1995. Harvesting of New Zealand native birds by Maori. Pages 54-59 in G. C. Grigg, P. T. Hale, and D. Lunney, editors. Conservation through sustainable use of wildlife. Center for Conservation Biology. University of Queensland, Brisbane, Australia.
Kitson, J. C. 2004. Harvest rate of sooty shearwaters (Puffinus griseus) by Rakiura Maori: a potential tool to monitor population trends? Wildlife Research 31, in press.
Klein, D. R., L. Moorehead, J. Kruse, and S. R. Braund. 1999. Contrasts in use and perceptions of biological data for caribou management. Wildlife Society Bulletin 27:488-498.
Kofinas, G. 2002. Community contributions to ecological monitoring: knowledge co-production in the U.S.-Canada Arctic borderlands. Pages 54-91 in I. Krupnik and D. Jolly, editors. The Earth is faster now: indigenous observations of Arctic environmental change. Arctic Research Consortium of the United States, Fairbanks, Alaska, USA.
Kofinas, G., P. Lyver, D. Russell, R. White, A. Nelson, and N. Flanders. 2004. Towards a protocol for monitoring of caribou body condition. Rangifer (Special Issue 14):43-52.
Krupnik, I., and D. Jolly, editors. 2002. The Earth is faster now: indigenous observations of Arctic environmental change. Arctic Research Consortium of the United States, Fairbanks, Alaska, USA.
Ludwig, D. 2001. The era of management is over. Ecosystems 4:758-764.
Lyver, P. 2002. The use of traditional environmental knowledge to guide sooty shearwater (Puffinus griseus) harvests by Rakiura Maori. Wildlife Society Bulletin 30:29-40.
Lyver, P., and A. Gunn. 2004. Calibration of hunters' impressions with female caribou body condition indices to predict probability of pregnancy. Arctic 57(3), in press.
Lyver, P., S. Hamilton, M. McKenzie, I. Dickson, M. Dooher, T. Broad, and H. Moller. 1998. Construction and reliability of an infra-red camera for examining nests in burrows. Conservation Advisory Science Notes 209:1-21.
Lyver, P., H. Moller, and C. Thompson. 1999. Changes in sooty shearwater Puffinus griseus chick production and harvest precede ENSO events. Marine Ecology Progress Series 188:237-248.
Mackinson, S. 2001. Integrating local and scientific knowledge: an example in fisheries science. Environmental Management 27:533-545
Manseau, M., J. Huot, and M. Crete. 1996. Effects of summer grazing by caribou on composition and productivity of vegetation: community and landscape level. Journal of Ecology 84:503-513.
Martin, P. S., and R.G. Klein, editors. 1984. Quaternary extinctions: a prehistoric revolution. University of Arizona Press, Tucson, Arizona, USA.
Moller, H. 1996. Customary use of indigenous wildlife; towards a bicultural approach to conserving New Zealand's biodiversity. Pages 89-125 in B. McFagen and P. Simpson, compilers. Biodiversity: papers from a seminar series on biodiversity. Department of Conservation, Wellington, New Zealand.
Moller, H. 2001. Co-management of a bicultural research project: a research provider's perspective. In M. Howard and H. Moller, editors. He Minenga Whakatü Hua o Te Ao [Sustaining the fruits of the land]: proceedings of a hui. (Murihiku Marae, 2000). Available online at: http://www.otago.ac.nz/Zoology/hui/Main/Talks2/HMoller.htm.
Moller, H. 2004. Are current harvests of seabirds sustainable? Acta Zoologica Sinica 50, in press.
Newman, J., and H. Moller. 2004. Use of Matauranga (Maori traditional knowledge) and science to guide a seabird harvest: getting the best of both worlds? In N. Kishigami and J. M. Savelle, editors. Indigenous use and management of marine resources. Senri Ethnological Studies Number 20. National Museum of Ethnology, Osaka, Japan, in press.
New Zealand Conservation Authority. 1997. Maori customary use of native birds, plants and other traditional materials. Interim report and discussion paper. NZCA, Wellington, New Zealand.
Posey, D. A. 1996. Traditional resource rights: international instruments for the protection and compensation for indigenous peoples and local communities. IUCN. Gland, Switzerland.
Porcupine Caribou Management Board. 2001. Porcupine Caribou Management Board 16th annual report 2001-2002. Whitehorse, Yukon, Canada.
Raffaelli, D., and H. Moller. 2000. Manipulative field experiments in animal ecology: do they promise more than they can deliver? Advances in Ecological Research 30:300-338.
Redford, K.H. 1992. The empty forest. Bioscience 42: 412-422.
Reidlinger, D., and F. Berkes. 2001. Contributions of traditional knowledge to understanding climate change in the Canadian Arctic. Polar Record 37:315-328.
Simpson, P. 2000. Dancing leaves; the story of New Zealand's cabbage tree, ti kouka. University of Canterbury Press, Christchurch, New Zealand.
Swezey, S. L., and R. F. Heizer. 1977. Ritual management of salmonid fish resources in California. Journal of California Anthropology 4:6-29.
Taiepa, T., P. Lyver, P. Horsley, J. Davis, M. Bragg, and H. Moller. 1997. Co-management of New Zealand's Conservation Estate by Maori and Pakeha: a review. Environmental Conservation 24:236-250.
Treseder, L., J. Honda-McNeil, M. Berkes, F. Berkes, J. Dragon, C. Notzke, T. Schramm, and R.J. Hudson. 1999. Northern Eden: community-based wildlife management in Canada. International Institute for Environment and Development, London, UK.
Walters, C. J., and C. S. Holling. 1990. Large-scale management experiments and learning by doing. Ecology 71:2060-2068.
Address of Correspondent:
Henrik Moller
Department of Zoology
University of Otago
P.O. Box 56
Dunedin, New Zealand
Phone: (643) 4797991
henrik.moller@stonebow.otago.ac.nz
![]()