|
|
|
Copyright © 2004 by the author(s). Published here under license by The Resilience Alliance.
Go to the pdf version of this article.
The following is the established format for referencing this article:
Blair, R. 2004. The effects of urban sprawl on birds at multiple levels of biological organization. Ecology and Society 9(5): 2. [online] URL: http://www.ecologyandsociety.org/vol9/iss5/art2/
A version of this article in which text, figures, tables, and appendices are separate files may be found by following this link.
Report, part of Special Feature on Urban Sprawl The Effects of Urban Sprawl on Birds at Multiple Levels of Biological Organization Robert Blair
University of Minnesota
- Abstract
- Introduction
- Methods
- Field sites: The gradients in California and Ohio
- Individual level: Nest predation and nesting success
- Species level: Bird distributions and densities
- Community level: Species richness, diversity, and life histories
- Landscape level: Landscape heterogeneity and structure of woody vegetation
- Continental level: Degree of faunal homogenization
- Results
- Field sites: The gradients in California and Ohio
- Individual level: Artificial nest predation and real nesting success
- Species level: Bird distributions and densities
- Community level: Species richness, diversity, and life histories
- Landscape level: Landscape heterogeneity and structure of woody vegetation
- Continental level: Degree of faunal homogenization
- Discussion
- Responses to this Article
- Acknowledgments
- Literature Cited
Urban sprawl affects the environment in myriad ways and at multiple levels of biological organization. In this paper I explore the effects of sprawl on native bird communities by comparing the occurrence of birds along gradients of urban land use in southwestern Ohio and northern California and by examining patterns at the individual, species, community, landscape, and continental levels. I do this by assessing the distribution and abundance of all bird species occupying sites of differing land-use intensity in Ohio and California. Additionally, I conducted predation experiments using artificial nests, tracked the nest fate of American Robins and Northern Cardinals, and assessed land cover in these sites. At the individual level, predation on artificial nests decreased with urbanization; however, this trend was not reflected in the nesting success of robins and cardinals, which did not increase with urbanization. At the species level, sprawl affected local patterns of extinction and invasion; the density of different species peaked at different levels of urbanization. At the community level, species richness and diversity peaked at moderate levels of urbanization, and the number of low-nesting species and of species with multiple broods increased with urbanization. The community-level results may reflect both the species-level patterns of local extinction and invasion as well as broader landscape-level patterns. At the landscape level, a linear combination of spatial heterogeneity and density of woody patches accurately predicted both species richness and Shannon Diversity. At the continental level, local extinction of endemic species, followed by the invasion of ubiquitous weedy species, leads to faunal homogenization between ecoregions.
KEY WORDS: avian community, California, Cardinalis cardinalis, extinction, faunal homogenization, invasion, landscape heterogeneity, nesting success, Ohio, predation, Turdus migratorius, urbanization.
Published: June 11, 2004
The ecology of sprawl
Urban sprawl affects the environment in myriad ways. Although urban expansion has resulted in the conversion of croplands, pastures, and forests into built environments on a massive scale (Alig and Healy 1987), little is known about the effects of urbanization at any level of biological organization from genes through landscapes. Ecologists have traditionally worked in relatively pristine environments (Cairns 1988) and, consequently, have failed to incorporate humans and their institutions as agents in the functioning of terrestrial ecosystems (McDonnell and Pickett 1990).
Although the ecology of urban areas has been an area of investigation for many years, including a paper that appeared in the first issue of Ecology (Huntington 1920), many researchers believed that, for the field of ecology to progress, studies had to occur in areas that were not subject to human pressures (Grimm et al. 2000). This view slowly changed during the 1990s with the publication of the work of Vitousek et al. (1997), which recognized the human domination of Earth's ecosystems, and with the establishment by the U.S. National Science Foundation of urban-based Long-Term Ecological Research sites in Baltimore, Maryland, and Phoenix, Arizona.
Growing interest in the ecological functioning of urban areas coincided with the realization that the United States is no longer a rural nation. In 1900, 40% of the U.S. population lived in urban areas, and 60% lived in rural areas. Today, 50% of the population of the United States lives in suburbs, and another 30% lives in cities (U.S. Census Bureau 2001). Migration to the suburbs is also increasing the extent of urban sprawl. Many urban areas are growing larger while the density of their human populations is simultaneously decreasing, with urban areas expanding at about twice the rate of the populations in many cities (Benfield et al. 1999, U.S. Department of Housing and Urban Development 2000). Smaller suburban communities actually occupy more land area than do major metropolitan areas, and migration to suburban communities will increase this difference (Katz and Bradley 1999). For example, of the 9224 km2 of urbanized land in Ohio, 7186 km2 are occupied by communities with fewer than 50,000 residents, whereas only 2038 km2 are occupied by more populous communities (U.S. Census Bureau 1990). As a result of the growth in urban settlement, approximately 5% of the United States is covered by urban areas, which is more than the area covered by national parks, state parks, and Nature Conservancy preserves combined (Stein et al. 2000, McKinney 2002).
Increased urban and suburban development and its subsequent sprawl can lead to huge conservation challenges (McKinney 2002). Urban development increases local extinction rates and the rates of loss of native species (Marzluff 2001). In addition, urban development is likely to lead to the replacement of native species by non-native species (Kowarik 1995, Blair 1996, Blair and Launer 1997, Blair 2001a,b). Although the severity of the disturbance caused by urban sprawl is similar to that caused by deforestation, it is more permanent, and the affected lands are less likely to revert to predisturbance conditions (McKinney 2002).
Urban ecosystems are diverse, and their landscapes do not conform to a simple dichotomy of urbanized vs. pristine. They do not fit into the simplistic paradigm of habitat and nonhabitat, because sprawl creates mosaics that are
more complex than pieces of cement and remnant native vegetation (Rebele 1994). Urban areas encompass a full range of manipulation from slightly altered open-space areas to highly developed commercial districts. Consequently, research in urban areas should consider the full range of urban and suburban land uses across a gradient of increasingly intense manipulation by humans (McDonnell and Pickett 1990, Pickett et al. 2001).
Most ecologists do not conduct research in human-dominated ecosystems and, consequently, are unfamiliar with the myriad terms that describe urban systems. Marzluff et al. (2001) attempted to bring some uniformity to this vocabulary and proposed that ecologists use the terms defined in Table 1. Although these terms are appropriate for large-grain studies, they are relatively gross for classifying individual land-use types. Consequently, I use these terms
generically in this paper but also refer to specific land uses. Finally, I wish to clarify the term "urbanization." Some authors refer to it as a process, with the concept of urbanization implying increasingly intense urban land use. Others refer to urbanization as a level of development. It is, in fact, both. Rarely does the use of a particular parcel of land progress sequentially along an urban gradient, although each parcel generally does fall somewhere along a gradient of land use intensity.
The effect of urbanization on bird communities has been an area of research since ~ 1950 (Marzluff 2001). Some researchers have compared the pre- and postdevelopment bird communities at a site (e.g., Graber and Graber 1963), whereas others have compared two sites with different levels of development (e.g., Beissinger and Osborne 1982). A few have attempted to examine a range of development intensities by comparing residential areas of different ages (e.g., Vale and Vale 1976) or by comparing areas with different land uses (e.g., Jokimaki and Suhonen 1993).
Recently, some researchers have turned to assessing bird communities across a range of urban land uses to examine the effects of spatial pattern (Hennings and Edge 2003), habitat fragmentation (Bolger et al. 1997), adjacent
landscapes (Clergeau et al. 2001), and scale (Hostetler 1999).
These studies suggest that it is important to examine the composition of the community and the distribution of individual birds as well as overall measures of the avian community such as species richness. Different groups of birds appear to be affected in different ways, and this has distinct conservation implications. Birds appear to fall into three broad groups: urban avoiders, suburban adaptable, and urban exploiters, each with different management needs (Blair 1996, McKinney 2002).
These studies also suggest that urbanization affects the heterogeneity of the landscape and, consequently, the distribution, abundance, and resources upon which birds depend. Typically, moderate development increases heterogeneity, the cover of ornamental vegetation, the availability of water sources, primary productivity, and the amount of edge between habitats. Extreme development, however, decreases heterogeneity and the availability of resources as they are permanently replaced with pavement and structures (Whitney and Adams 1980, Godron and Forman 1983, Mooney and Gulmon 1983, Rudnicky and McDonnell 1989).
Urbanization leaves a clear signature on the distribution patterns of birds. The intent of this paper is to examine the effects of urbanization on birds at multiple levels of biological organization by assessing (1) the success of nesting
individuals, (2) the density and distribution of species, (3) the diversity and species richness of communities, and (4) the distribution of communities across the landscape and continent of North America. Here I present previously reported and new research at all of these levels of biological organization and speculate about mechanistic links between levels to provide a more comprehensive view of how urbanization alters bird distributions.
The sites in this study spanned two full gradients of urban development that reflect the typical urban-suburban matrix that results from sprawl. The sites ranged from relatively undisturbed to highly developed and included biological preserves, open-space reserves, golf courses, residential neighborhoods, an office park (California) or apartment complexes (Ohio), and business districts. I considered the office park in California and the apartment complexes in Ohio to be comparable because of their similarity in scale and in the layout of both buildings and plantings. I made this substitution because no office parks exist in the vicinity of the other sites in Ohio. The area sampled within each site was approximately 16 ha. These gradients of sites are located in two ecoregions that the U.S. Geological Survey identifies as coastal chaparral forest shrub in California and eastern broadleaf forest in Ohio
(Bailey 1994).
In California, the six sites were located within a circle with a 3-km radius centered at Stanford University near Palo Alto (37° 20' N, 122° 15' W, with a typical elevation of < 100 m). The California sites are shown in Fig. 1. In Ohio, the six sites were within a 10-km radius of Miami University, Oxford (39° 30' N, 84° 45' W, typical elevation ~ 290 m). The Ohio sites are shown in Fig. 2. I selected sites within each ecoregion that were as ecologically similar to one another as possible prior to development according to historical maps and reference materials (see Blair 1996 and Porter et al. 2001 for more detailed descriptions of the sites).
Because urban and suburban sprawl is an exceedingly complex amalgamation of factors, it is often difficult to array sites along a single axis of urbanization (McDonnell et al. 1993). However, for this study, I used land cover as a surrogate measure of urbanization. At each study site, I calculated the area covered by buildings, pavement, lawn, grassland, and trees or shrubs in a 50-m radius centered at each of 16 points used for the bird surveys described below. I then converted the areas to percentage of site covered. In California, I estimated the cover of each land-use type from recent aerial photographs provided by Stanford University, the City of Palo Alto, and Stanford University's Jasper Ridge Biological Preserve. In Ohio, I used orthodigitally corrected land-use maps from the City of Oxford
and aerial photographs from the Department of Geography at Miami University.
I used Principal Component Analysis (PCA) in PCOrd (McCune and Mefford 1999) to assign the sites a value that represented their level of urbanization on the gradient. Specifically, I used the estimates of percent cover of buildings, pavement, lawn, grassland, and trees or shrubs in a PCA and assigned each site the score from the first axis of the analysis as its level of urbanization.
To examine the effects of urbanization on potential predation pressure on nests, a field assistant and I deployed artificial nests in each of the Ohio sites during June of 1999 and 2000. Each nest contained two Zebra Finch (Poephila guttata) eggs and one hand-formed plasticine egg and was placed 0–2 m above the ground. We left the nests in position for 12 days to simulate the nesting period of most passerines (Gill 1995). We classified any nest in which an egg had been broken or removed as depredated. We classified nests found in their original state as undisturbed (for details, see Reale 2002).
To examine the effects of urbanization on actual nests, we followed the nesting attempts of American Robins (Turdus migratorius) and Northern Cardinals (Cardinalis cardinalis), which typically nest less than 3 m above the ground (Harrison 1975). We selected these two species because they are found at all sites along the gradient of
urbanization and their nesting habits could be mimicked with artificial nests. In May and June 2000, a field assistant and I located nests of robins and cardinals at all the sites in Ohio. After we found the nests, we used a convex mirror mounted on a 10-m expandable pole (Parker 1972) to examine the contents while minimizing contact with the nests. We located all the nests during the building, egg-laying, or early incubation stages and monitored them every three to four days during incubation and after hatching to determine fledging success. For each nest, we recorded the number of eggs laid, hatched, and fledged as well as any predation or egg loss.
To compare artificial nests to real nests, we constructed a logistic regression model with site, nest type, and the interaction between site and nest type as independent variables. This model tested three hypotheses simultaneously: (1) artificial and real nests have the same probability of being predated across sites, i.e., the slopes of the predicted probability of predation across sites for artificial and real nests are the same; (2) artificial and real nests have the same intercepts but possibly different slopes, i.e., urbanization affects either real or artificial nests to a greater degree; and (3) the relative probability of predation within sites is the same, i.e., the probability of predation is not related to the level of urbanization of a site (Oris and Bailer 1997). We used the Wald statistic (W) for all three model factors, i.e., site, species, and interaction, to test these hypotheses. We considered factors as significant if p < 0.05.
I used variable circular plots to estimate the densities of all perching or singing birds during peak breeding season in both California and Ohio. I established 16 survey points within each site in an approximately 4 x 4 matrix where each
point was at least 100 m from its nearest neighbor (Reynolds et. al 1980). In the preserve and open-space reserve in Ohio, I located sampling points along foot trails instead of in a 4 x 4 matrix at the request of the areas' managers.
To assess the validity of this method, an undergraduate and I compared sampling along trails and in a 4 x 4 matrix in the nature preserve in 2002. We found no significant differences between these two methods in determining the
densities of any of the species at the site (R. Blair, unpublished data).
In California, a field assistant and I surveyed points a total of eight times in June and July 1992, and four times in June 1993. The open-space site was visited only six times in 1992 because it burned on 10 July. In Ohio, we surveyed
points a total of eight times in June and July 1996, and four times in June 1997. Surveys began at dawn and continued until we had covered all 16 points at a site, which took approximately 2 h. We visited each point for 5 min. This method resulted in an estimate of the absolute density (birds/ha) of all the species within each site (for details on method, see Blair 1996).
To assess the role of local extinction and invasion, I examined the best representatives of the communities that existed in each ecoregion prior to European settlement, i.e., those species found in the preserves, which are also known as the minimally disturbed standard (Karr and Chu 1998). I assumed that local extinction along the urban
gradient had occurred if a given species was found in relatively undisturbed sites but was not found at more urbanized sites. I assumed that local invasion had occurred if a species was found only at more urbanized sites along the
gradient. These definitions are not mutually exclusive. For example, House Finches (Carpodacus erythrinus) do not occur at either end of the gradient in California but do occur at sites with intermediate levels of urbanization.
This would seem to indicate that the species invades sites with low levels of urbanization but is then extirpated from such sites as their level of urbanization increases.
I calculated species diversity using the Shannon Index (Shannon and Weaver 1949, Magurran 1988). I defined species richness for each site as the total number of species recorded, because effort and area covered were approximately
equal at all sites. To examine life history characteristics, I classified each species according to (1) breeding strategy, i.e., (single brood, double broods, or multiple broods per season; (2) nesting height, i.e., low (0–3 m), mid-height
(3–6 m), and high (> 6 m); and (3) nest type, i.e., open or cavity, based on Ehrlich et al. (1988). I analyzed the community and life history measures with linear regression to determine if they were significantly correlated with the
degree of urbanization.
To determine if landscape heterogeneity was related to the Shannon Diversity and species richness of birds, I calculated percent dissimilarity (PD) of all the woody species sampled in circular plots with a radius of 0.04 ha or 11.3 m centered at all the sampling points in Ohio (Collins 1992, Porter et al. 2001). For simplicity, I excluded all plots that did not have any woody vegetation; these included six sites in the business district, six in the apartment complexes, three in the residential area, seven on the golf course, one in the open-space recreational area, and one in the nature preserve. I used Kruskall Wallis one-way ANOVA to analyze variation in PD among sites. I then used linear regression to determine if bird species richness and Shannon Diversity were significantly correlated with landscape heterogeneity
as represented by percent dissimilarity of woody species. I also calculated the number of patches of woody vegetation per hectare, the percentage of canopy cover per hectare, and the average patch size. I used these factors in a forward
stepwise multiple linear regression to model both species richness and Shannon Diversity as factors of landscape variables (see Porter et al. 2001 for details).
To determine if increasing urbanization led to a more homogeneous fauna, I compared sites within land use type between ecoregions, e.g., the golf course in California with the golf course in Ohio, by using a Pearson correlation on Jaccard's Index of Similarity, which measures percent overlap in species composition between two sites (Magurran 1988). Jaccard's Index ranges from 0, indicating no species overlap, to 1, indicating complete species overlap.
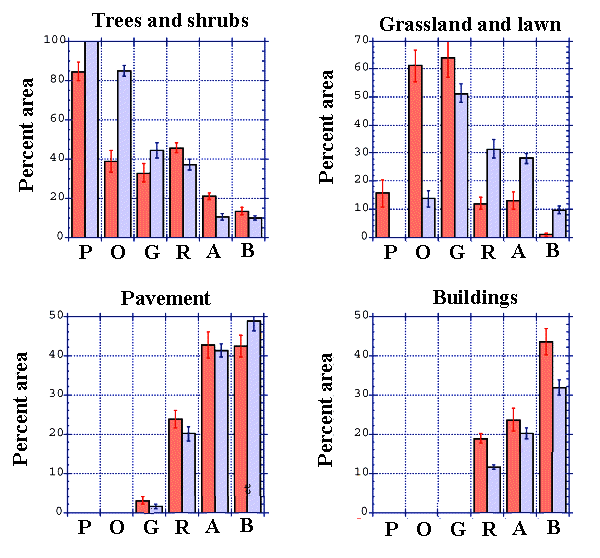
In California, the first axis of the Principal Component Analysis (PCA) on land cover explained 73.9% of the variance in land cover among sites. Scores along the axis were as follows: nature preserve (-1.65), open space (-1.65), golf course (-1.49), residential district (0.48), research park (1.63), and business district (2.68). In Ohio, the first axis of the PCA for the sites explained 70.1% of the variance in land cover among sites. Scores along the axis were as follows: nature preserve (-1.99), open space (-1.70), golf course (-0.84), residential district (0.30), apartment complex (1.75), and business district (2.48). The rank order of sites in both California and Ohio from most natural to most urban coincided with surveys of researchers familiar with the sites based on their perceived level of urbanization of the sites (Delphi surveys; for details, see Blair 1996, Gering and Blair 1999). The rank order was confirmed further by the unimodal
distributions of land cover types (Fig. 3).
Of the 170 artificial nests, 119 were depredated. Of real nests, we discovered 85 in the five most urban sites: 51 occupied by robins and 34 occupied by cardinals. There were too few nests in the preserve to include them in this analysis. We recorded a total of 36 predation events. Fifty-three nests successfully fledged at least one offspring. Predation did not always result in nest destruction, and some nests were successful despite the loss of one or more eggs. There were no significant differences in the probability of predation between robins and cardinals at any nesting stage (W = 1.20, p = 0.23). Consequently, we pooled data for the two species to compare with artificial nests. The average height of nests in a site was 1–2.5 m for robins and 1–2 m for cardinals (Reale 2002). Consequently, the height of the artificial nests approximated the distribution of the real nests.
For the comparison of predation on artificial nests and on real nests during the entire nesting period, site was a significant predictor of predation (W = -2.05, p = 0.04) as was nest type (W = 3.44, p = 0.001). The probability of predation decreased with increasing urbanization. Further, the significance of nest type indicated that predation on the two nest types was not equal; artificial nests had higher probabilities of predation than real nests. Additionally, there was no significant interaction between nest type and site (W = 0.79, p = 0.78), which indicated that predation on artificial nests with respect to urbanization mirrored that on real nests, although to a greater degree. The decrease in predation with increasing urbanization did not increase the nesting success of robins and cardinals. Site was not a significant predictor of nest fate for robins (df = 1, W = 0.66, p = 0.42) or cardinals (df = 1, W = 0.002, p = 0.96). This implies that the reasons for nest failure shift from
predation in more natural sites to other reasons such as nest abandonment in more urbanized sites.
The densities of all bird species varied across the urban gradients (Tables 2 and 3). Of the 40 bird species we recorded in California, 31 had continuous unimodal distributions of abundance across the urban gradient within the limits of one standard error of their estimated daily densities. Of the 44 species encountered in Ohio, 38 had unimodal distributions within the limits of one standard error.
Woodland species, the minimally disturbed standard, displayed a strong pattern of local extinction in both ecoregions. Most of the woodland species dropped out along the gradient as the sites became more urbanized. Only three of the
21 bird species present in the preserve in California also occurred in the business district. Only one of the 17 bird species present in the preserve in Ohio also occurred in the business district (Table 4). These patterns were
also highly nested, i.e., species in relatively depauperate sites were subsets of those in more species-rich sites. Once a woodland species dropped out at a less urbanized site, for example, it rarely reappeared at a more urbanized
site along the gradient.
The pattern of invasion by bird species along the gradients was strong as well. In California, 19 species were categorized as local invaders because they were not present at the preserve but were present at more urbanized sites. In Ohio, 27 species were identified as local invaders. However, local invaders were not necessarily safe from local extinction as a result of urbanization. Of the local invaders, 14 species in California and 20 species in Ohio did not occur in the most urbanized sites. These species exploited suburban conditions but were not able to tolerate
the most urban conditions.
Species richness in California was unimodal and significantly related to urbanization in a quadratic multiple linear regression (n = 6, R2 = 0.868, p < 0.01). The relationship between species richness and urbanization in Ohio was marginally significant (n = 6, R2 = 0.811, p = 0.0820). Shannon Diversity also was unimodal and significantly related to urbanization (California: n = 6, R2 = 0.983, p < 0.005; Ohio: n = 6, R2 = 0.860, p = 0.053). This implies that species
richness and diversity actually increase with moderate levels of urbanization and then decrease at higher levels of urbanization.
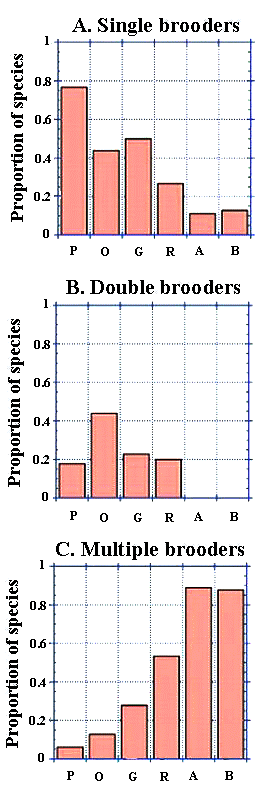
I documented 43 species across all six sites of the urban gradient in Ohio but excluded Brown-headed Cowbird (Molothrus ater) from the community-level guild analysis because of its unique status as a brood parasite. Of the 42 remaining species, 29% (n = 12) species were high nesting, 33% (n = 14) were mid-height nesting, and 37% (n = 16) were low nesting. Tree-nesting species accounted for 55% (n = 23), and shrub/ground-nesting species made up 31% (n = 13). An additional 14% (n = 6) had some other nesting location. Fifty-five percent of the species were single brooding (n = 23), 24% were double brooding (n = 10), and 21% were multiple brooding (n = 9). Nesting height was inversely correlated with the degree of urbanization at a marginal level of significance (n = 6, R2 = 0.96, p = 0.06; Fig. 4). The brood strategy of species differed significantly across sites; the number of broods increased as the level of urbanization increased (n = 6, R2 = 0.93, p < 0.01; Fig. 5).
In Ohio, spatial heterogeneity, as measured by percent dissimilarity of woody species, varied significantly among sites (X2 = 308.09, df = 5, p < 0.001) and peaked in sites with moderate urbanization. However, spatial heterogeneity was not significantly correlated with either species richness (n = 6, R2 = 0.017, p = 0.80) or with Shannon Diversity (n = 6, R2 = 0.062, p = 0.64). A linear regression model using both percent dissimilarity of woody vegetation and the average number of canopy patches per hectare did significantly predict both species richness (n = 6, R2 = 0.926, p = 0.02) and Shannon Diversity (n = 6, R2 = 0.940, p = 0.01; Table 5).
The similarity of the species composition of birds, which is assessed with Jaccard's Index, between the sites in California and Ohio was 0.06 for the biological preserves, 0.03 for the open-space reserves, 0.11 for the golf courses, 0.15 for the residential areas, 0.22 for the office park/apartment complexes, and 0.15 for the business districts. Similarity of species composition was significantly correlated with the degree of urbanization in both California (n = 6, R2 = 0.660, p = 0.05) and Ohio (n = 6, R2 = 0.751, p = 0.02).
These results suggest that urban sprawl affects birds at multiple levels of biological organization, from individuals on a local scale to communities across a continent. What occurs at one level of biological organization may not seem linked to what occurs at another, but connections can be made between such phenomena as local extirpation because of alterations to the landscape and continental homogenization of avifauna. Although the research presented here does not address every effect and every mechanism of how sprawl alters bird distributions, it does begin to suggest a comprehensive framework for addressing the impacts of sprawl and proposing means for conserving native species.
At the individual level, urbanization altered the probability of predation on both real and artificial nests. In both of the experiments, predation decreased with increasing urbanization, although artificial nests experienced significantly
higher probabilities of predation than did real nests across the urban gradient. This work apparently contradicts that of other researchers in two ways: (1) artificial nests can serve as surrogates for real nests, and (2) predation decreases
with increased urbanization.
Several researchers contend that predation rates on artificial nests do not reflect predation rates on real nests (George 1987, Martin 1987, Storaas 1988, Reitsma et al. 1990, Roper 1992). We found that the probability of predation is notably higher for artificial nests than real nests, but the probabilities of predation for both artificial and real nests decline as urbanization increases. In other words, artificial nests can be used as a surrogate measure of relative predation pressure on real nests under different intensities of urbanization. However, I did not find a link between nesting success and predation for cardinals and robins in Ohio. Traditionally, ornithologists have viewed nest predation as the driving factor in nesting success (Ricklefs 1969, Emlen 1974, Martin 1988). In this work, nesting success was more strongly affected by a variety of other factors such as nest abandonment and the death of nestlings in highly urban areas. Consequently, even though the probability of nest predation decreased with urbanization, I did not detect a concurrent increase in nesting success. It should be noted that our focal species are somewhat aggressive and opportunistic. Therefore, robins and cardinals may not represent the response of species that are more sensitive to human development. However, these two species are extremely useful because they can be found in almost every land-use type, which allows researchers to conclude that differences in the behavior of individual birds may be attributed to land use and not differences in species.
These findings also apparently contradict those of researchers who have found that nest predation increases with urbanization. For example, Jokimaki and Huhta (2000) found that predation on artificial nests stocked with quail eggs
was higher in urban parks than in neighboring woodlands in Finland. The latter findings were supported by Sorace (2002), who estimated abundances of pests such as pigeons, starlings, rats, and house mice in open areas in urban parks and neighboring woodlands and found that pests were more abundant in the urban parks. These findings might not be incongruous with those presented here. I surveyed different land-use types as opposed to woodlands with different adjacent land uses. The idea that adjacent land use affects predator populations is supported by Thorington and Bowman (2003), who placed artificial nests stocked with quail eggs in natural oak scrub patches near residential areas of varying human density. They found that predation was higher in the patches with higher housing densities and cautioned that predation pressures may vary within single land-use types.
As mentioned previously, predation may be only one piece of the mechanism driving nesting success. I found that predation decreased with increasing urbanization but that the nesting success of robins and cardinals did not necessarily increase. This may be because of other factors, including the survival of adults in the nesting period, myriad causes of nest abandonment, and erratic food supplies. Antonov and Atanasova (2003) compared the reproduction of magpies (Pica pica) in rural and urban areas of Bulgaria. They found that urban magpies had lower predation but also suffered brood reduction, which resulted in the production of fewer fledglings per pair than for rural magpies.
At the species level, I found that the distribution of individual species varied predictably across the urban gradient in the sense that the densities of the species displayed unimodal distributions, which peaked at one level of development and decreased with either greater or lesser urbanization. Some species were urban avoiders with their highest densities at the most natural sites, e.g., the Kentucky Warbler (Oporornis formosus) in Ohio. Most species were able to adapt to the suburbs, with densities peaking at intermediate levels of development, e.g., the American Robin in both California and Ohio. Some species were urban exploiters whose highest densities were found at the most urban sites, e.g., the European Starling (Sturnus vulgarus) in Ohio (Blair 1996, McKinney 2002).
This ebb and flow of species densities across the urban gradient suggests that some individual species disappear with urbanization, whereas other species invade in response to the environmental changes associated with development. It is possible that this pattern of extinction and colonization is influenced by the changes in predation rates and the different causes of nest failure that occur with urbanization. From a conservation standpoint, it is important to note that almost all of the woodland species that existed in intact forests in California and Ohio did not occur in the most urban sites and that, on the whole, woodland species gradually dropped out as land use became more urban. For instance, the golf courses in California and Ohio supported only 50% of the woodland species that presumably comprised the native bird community prior to their construction.
Urbanization induces clear changes in the bird community as a whole. Species richness and Shannon Diversity actually peaked at intermediate levels of urbanization and decreased with either more or less development. This phenomenon creates the interesting conundrum that the golf courses in California and Ohio were the most species-rich sites along their respective urban gradients and also contained only half of the woodland species. This phenomenon can also be linked to local extinction and invasion. The slight development of a site can create more heterogeneous conditions that support more species of birds, but severe development apparently curtails the resources necessary for even the suburban-adaptable invaders.
Changes in the bird community were also documented in the nesting habits of the birds across the gradient. The height of trees in urban areas is generally lower than in less developed areas (Porter et al. 2001) and, consequently,
urban areas offer fewer nesting options (Reale 2002). This was reflected in the study systems; species with high nests dropped out completely from the more urban end of the gradient, whereas the density of mid-height- and low-nesting
species increased. Even more interesting was the change in the brooding strategy of the community. The proportion of single-brooding species in the community declined with increasing urbanization, whereas the proportion of multiple-brooding species steadily increased. This result may seem counterintuitive given the finding that predation decreased with urbanization, but it is tempered by the fact that the nesting success of robins and cardinals did not change across
the urban gradient. Apparently, other nesting pressures are replacing predation with increasing urbanization. I found many other causes of mortality, such as nest abandonment and nestling death, that could not be explained by predation. In part, this shift in nesting pressure may also explain the shift in the brooding strategy of the community. An interesting area of future research would be to examine whether single species, such as cardinals, also display this shift in brooding strategy with urbanization.
At the landscape level, I found that landscape heterogeneity as measured by woody vegetation varied across the urban gradient and peaked at intermediate levels of development. Because species richness and Shannon Diversity also peaked at intermediate levels of development, I expected landscape heterogeneity to be another force driving community composition. However, heterogeneity and richness were not directly correlated. To devise a model that accurately predicted the number of species or Shannon Diversity of a site, I had to include the number of canopy patches per hectare and the heterogeneity of the woody vegetation. Surprisingly, with this two-variable model, species richness or diversity was positively correlated with heterogeneity and negatively correlated with the number of canopy patches. Apparently, bird diversity increases when a variety of woody vegetation types is present in an area, provided that the extent of fragmentation in each of those types is relatively low.
The patterns of interaction between the heterogeneity of woody vegetation and the number of canopy patches provide further insight into how urbanization may lead to local extinction and invasion simultaneously. In general, the spatial heterogeneity of vegetation is associated with species richness (Ricklefs and Schluter 1993). However, the association between heterogeneity and species richness is tempered in urban systems, in which it appears that the fragmentation of the canopy results in the loss of woodland species. Once again, local invasion, facilitated at
the landscape level by increased heterogeneity, is tempered by local extinction in response to fragmentation of the landscape.
At the continental level, I saw distinct homogenization of the avifauna with urbanization. The overlap in the bird communities in California and Ohio increased from approximately 5% in the least developed sites to approximately 20% in the most urbanized sites. Although a leap from 5 to 20% may not sound alarming, it does highlight a global conservation challenge. In many instances, local extinction of endemic species is followed by local invasion by ubiquitous species. Apparently, it is not a serendipitous circumstance that House Sparrows (Passer
domesticus) can be found begging for french fries outside McDonald's restaurants anywhere in the world.
Although this paper covers a range of ideas on how urbanization alters birds at multiple levels of biological organization from reduction in individual nesting success to continental homogenization of the avifauna, it is by no means comprehensive. Instead, I have presented an overview of the work that I have conducted at different levels of biological organization and suggested ways in which those levels may be linked through extinction and invasion. The idea of extinction followed by invasion may also prove to be a useful conceptual model for future research in urban systems. Although I have focused on birds, I suspect that much of this work may be generally applicable to other animal taxa (see also Blair 1999, Blair 2001b). This model of extinction and invasion may be linked to gaps in
the spectrum of body masses exhibited in the community (Allen et al. 1999, Hostetler 1999). Allen et al. (1999) found that body mass patterns were correlated to invasion and extinction in human-transformed ecosystems at large scales,
e.g., the Florida Everglades. The results presented here may hint at why these correlations exist.
Urban-avoiding and urban-exploiting species share some common traits that emerge at all levels of organization. Examining these traits collectively demonstrates that the processes that affect individuals may play out on a continental scale, changing bird communities in somewhat predictable ways (Fig. 6).
Future research on birds and urbanization should examine further the mechanisms that bring about these changes. Additionally, it should examine the nuances of these patterns. All birds are not the same, and it may be useful to examine neotropical migrants separately from short-distance migrants and year-round residents (Hennings and Edge 2003). Likewise, not all land use is the same. It would be worthwhile to dissect the differences of single land-use types
to discover why some variations are better at conserving native species than others (Thorington and Bowman 2003). Finally, it would be interesting to examine patterns of community structuring that occur at larger scales.
Table 1. Standardized terms that describe major points along a gradient or urbanization. These terms are most meaningful at relatively large ( > 1 km2) scales (Marzluff et al 2001).
![]()
Term
![]()
Wildland
Rural/exurban
Suburban
Urban
![]()
Birds in urban systems
Fig. 1. Aerial views of sites in California. Each covers approximately 50 ha.

Fig. 2. Aerial views of sites in Ohio. Each covers approximately 50 ha.

Fig. 3. Percent area (± SE, n = 16 points) covered by trees and shrubs, grassland and lawn, pavement, and buildings in California (darker) and Ohio (lighter). The x-axis reads as follows: P = preserve, O = open space, G = golf course, R = residential, A = apartment complexes/research park, and B = business district.
Individual level: Artificial nest predation and real nesting success
Table 2. Distribution and abundance of summer resident birds in Palo Alto, California. As portrayed in the legends at the beginning and end of the table, line widths represent ranges of numbers of birds per hectare. These ranges are displayed graphically to illustrate the ebb and flow of densities across the gradient for each species. Nomenclature follows the American Ornithologists' Union (1998).

Table 3. Distribution and abundance of summer resident birds in Oxford, Ohio. As portrayed in the legends at the beginning and end of the table, line widths represent ranges of numbers of birds per hectare. These ranges are displayed graphically to illustrate the ebb and flow of densities across the gradient for each species. Nomenclature follows the American Ornithologists' Union (1998).

Table 4. Loss of the original woodland species across the urban gradients in California and Ohio. A plus sign (+) indicates presence. Nomenclature follows the American Ornithologists' Union (1998).
![]()
Bird species
![]()
California ecoregion
Hutton's Vireo
Western Wood-Pewee
Steller's Jay
Wrentit
Dark-eyed Junco
Ash-throated Flycatcher
Blue-gray Gnatcatcher
Cliff Swallow
White-breasted Nuthatch
California Quail
Violet-green Swallow
Spotted Towhee
Bewick's Wren
Lesser Goldfinch
Plain Titmouse
California Towhee
Bushtit
Chestnut-backed Chickadee
Scrub Jay
Anna's Hummingbird
Mourning Dove
Ohio ecoregion
Acadian Flycatcher
American Crow
Great Crested Flycatcher
Kentucky Warbler
Red-eyed Vireo
Ovenbird
White-breasted Nuthatch
Northern Parula
Indigo Bunting
Tufted Titmouse
Blue-gray Gnatcatcher
Red-bellied Woodpecker
Downy Woodpecker
Eastern Wood-pewee
Brown-headed Cowbird
Carolina Chickadee
Northern Cardinal
![]()
Fig. 4. The proportion of species present in each site presented in order of increasing urbanization for (A) high-nesting, (B) mid-height-nesting, and (C) low-nesting species. The x-axis reads as follows: P = preserve, O = open space, G = golf course, R = residential, A = apartment complexes, and B = business district.

Fig. 5. The proportion of species present in each site presented in order of increasing urbanization for (A) single brooders, (B) double brooders, and (C) multiple brooders. The x-axis reads as follows: P = preserve, O = open space, G = golf course, R = residential, A = apartment complexes, and B = business district.
Landscape level: Landscape heterogeneity and structure of woody vegetation
Table 5. Regression estimates, probability, proportion of variance explained, and F-values for multiple linear regression equations of species richness and Shannon Diversity.
![]()
![]()
Term
Intercept
Average percent dissimilarity
Patches/ha
![]()
![]()
Term
Intercept
Average percent dissimilarity
Patches/ha
![]()
Continental level: Degree of faunal homogenization
Fig. 6. Conceptual model of how local extinction and invasion are connected by urbanization at multiple levels of biological organization.

Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a comment, follow this link. To read comments already accepted, follow this link.
The work presented here is that of a number of people. For simplicity, I have often written in the first person singular, but, in actuality it has been a collective effort by many people over the past decade. Alistair Hobday, Charlie Quinn, Jay Mager, John Minturn, Tom Minturn, and Brian Forschner were field assistants on various facets of the research. Jon Gering and Jospeh Reale conducted field experiments and wrote various manuscripts as graduate students. Eric Porter assembled the information on woody vegetation as part of his postdoctoral studies. Paul Ehrlich, Dave Berg, Mercy Ebbert, Mike Vanni, Bruce Steinly, and Nancy Solomon have provided me with many intellectual insights over the years. Finally, I am particularly indebted to Erica Fleishman for both intellectual and editorial prodding.
Alig, R. J., and R. G. Healy. 1987. Urban and built-up land area changes in the United States: an empirical investigation of determinants. Land Economics 63:215-226.
Allen, C. R., E. A. Forys, and C. S. Holling. 1999. Body mass patterns predict invasions and extinctions in transforming landscapes. Ecosystems 2:114-121.
American Ornithologists' Union. 1998. Check-list of North American birds: the species of birds of North America from the Arctic through Panama, including the West Indies and Hawaiian Islands. AOU, Washington, D.C., USA.
Antonov, A., and D. Atanasova. 2003. Small-scale differences in the breeding ecology of urban and rural Magpies Pica pica. Ornis Fennica 80:21-30.
Bailey, R. G., P. E. Avers, T. King, and W. H. McNab, editors. 1994. Ecoregions and subregions of the United States. U.S. Geological Survey, Washington, D.C., USA.
Beissinger, S. R., and D. R. Osborne. 1982. Effects of urbanization on avian community organization. Condor 84:75-83.
Benfield, F. K., M. D. Raimi, and D. D. Chen. 1999. Once there were greenfields. Natural Resources Defense Council, New York, New York, USA.
Blair, R. B. 1996. Land use and avian species diversity along an urban gradient. Ecological Applications 6(2):506-519.
Blair, R. B. 2001a. Birds and butterflies along urban gradients in two ecoregions of the United States: is urbanization creating a homogeneous fauna? Pages 33-56 in J. L. Lockwood and M. L. McKinney, editors. Biotic homogenization. Kluwer Academic/Plenum, New York, New York, USA.
Blair, R. B. 2001b. Creating a homogeneous avifauna. Pages 459-486 in J. M. Marzluff, R. Bowman, and R. Donnelly, editors. Avian ecology and conservation in an urbanizing world. Kluwer Academic, Norwell, Massachusetts, USA.
Blair, R. B., and A. E. Launer. 1997. Butterfly diversity and human land use: species assemblages along an urban gradient. Biological Conservation 80:113-125.
Bolger, D. T., T. A. Scott, and J. T. Rotenberry. 1997. Breeding bird abundance in an urbanizing landscape in coastal southern California. Conservation Biology 11:406-421.
Cairns, J. 1988. Restoration ecology: the new frontier. Pages 2-11 in J. Cairns, editor. Rehabilitating damaged ecosystems. CRC Press, Boca Raton, Florida, USA.
Clergeau, P., J. Jokimaki, and J. L. Savard. 2001. Are urban bird communities influenced by the bird diversity of adjacent landscapes? Journal of Applied Ecology 38:1122-1134.
Collins, S. L. 1992. Fire frequency and community heterogeneity in tallgrass prairie vegetation. Ecology 73(6):2001-2006.
Emlen, J. T. 1974. An urban bird community in Tucson, Arizona: derivation, structure, regulation. Condor 76:184-197.
Ehrlich, P. R., D. S. Dobkin, and D. Wheye. 1988. The birder's handbook: a field guide to the natural history of North American birds. Simon and Schuster, New York, New York, USA.
George, T. L. 1987. Greater land bird densities on island vs. mainland; relation to nest predation level. Ecology 68:1393-1400.
Gering, J. C., and R. B. Blair. 1999. Predation on artificial bird nests along an urban gradient: predatory risk or relaxation in urban environments? Ecography 22: 532-541.
Gill, F. B. 1995. Ornithology. W. H. Freeman, New York, New York, USA.
Godron, M., and R. T. Forman. 1983. Landscape modification and changing ecological characteristics. Pages 12-28 in H. A. Mooney and M. Godron, editors. Disturbance and ecosystems. Springer-Verlag, New York, New York, USA.
Graber, R. R., and J. W. Graber. 1963. A comparative study of bird populations in Illinois, 1906-1909 and 1956-1958. Illinois Natural History Survey Bulletin 28:377-527.
Grimm, N. B., J. M. Grove, S. T. A. Picket, and C. L. Redman. 2000. Integrated approaches to long-term studies of urban ecological systems. BioScience 50:571-584.
Harrison, H. H. 1975. A field guide to birds' nests. Houghton Mifflin, Boston, Massachusetts, USA.
Hennings, L. A., and W. D. Edge. 2003. Riparian bird community structure in Portland, Oregon: habitat, urbanization, and spatial scale patterns. Condor 105:288-302.
Hostetler, M. 1999. Scale, birds, and human decisions: a potential for integrative research in urban ecosystems. Landscape and Urban Planning 45(1):15-19.
Huntington, E. 1920. The control of pneumonia and influenza by the weather. Ecology 1(1):6-23.
Jokimaki, J., and E. Huhta. 2002. Artificial nest predation and abundance of birds along an urban gradient. Condor 102(4): 838-847.
Jokimaki, J., and J. Suhonen. 1993. Effects of urbanization on the breeding bird species richness in Finland: a biogeographical comparison. Ornis Fennica 70:71-77.
Karr, J. R., and E. W. Chu. 1998. Restoring life in running waters: better biological monitoring. Island Press, Washington, D.C., USA.
Katz, B., and J. Bradley. 1999. Divided we sprawl. Atlantic Monthly 284(6):26-42.
Kowarik, I. 1995. On the role of alien species in urban flora and vegetation. Pages 85-103 in P. K. Pysek, M. Prach, M. Rejmanek, and P. M. Wade, editors. Plant invasions: general aspects and special problems. SPB Academic, Amsterdam, The Netherlands.
Magurran, A. E. 1988. Ecological diversity and its measurement. Princeton University Press, Princeton, New Jersey, USA.
Martin, T. E. 1987. Artificial nest experiments: effects of nest appearance and type of predator. Condor 89:925-928
Martin, T. E. 1988. Habitat and area effects on forest bird assemblages: is nest predation an influence? Ecology 69:74-84.
Marzluff, J. M. 2001. Worldwide urbanization and its effects on birds. Pages 19-47 in J. M. Marzluff, R. Bowman, and R. Donnelly, editors. Avian ecology in an urbanizing world. Kluwer Academic, Norwell, Massachusetts, USA.
Marzluff, J. M., R. Bowman, and R. Donnelly. 2001. A historical perspective on urban bird research: trends, terms, and approaches. Pages 1-18 in J. M. Marzluff, R. Bowman, and R. Donnelly, editors. Avian ecology in an urbanizing world. Kluwer Academic, Norwell, Massachusetts, USA.
McCune, B., and J. B. Grace. 2002. Analysis of ecological communities. MJM Software Design, Gleneden Beach, Oregon, USA.
McDonnell, M. J., and S. T. A. Pickett. 1990. Ecosystem structure and function along urban-rural gradients: an unexploited opportunity for ecologists. Ecology 71:1232-1237.
McKinney, M. L. 2002. Urbanization, biodiversity, and conservation. BioScience 52(10):883-890.
Mooney, H. A., and S. L. Gulmon. 1983. The determinants of plant productivity—natural versus man-modified communities. Pages 146 - 158. in H. A. Mooney and M. Godron, editors. Disturbance and ecosystems. Springer-Verlag, New York, New York, USA.
Oris, J. T., and A. J. Bailer. 1997. Equivalence of concentration-response relationships in aquatic toxicology studies: testing and implications for potency estimation. Setac 16:2204-2209.
Parker, J. W. 1972. A mirror and pole device for examining high nests. Bird-Banding 43:216-218.
Pickett, S. T. A., M. L. Cadenasso, J. M. Grove, C. H. Nilon, R. V. Pouyat, W. C. Zipperer, and R. Costanza. 2001. Urban ecological systems: linking terrestrial ecological, physical, and socioeconomic components of metropolitan areas. Annual Review of Ecology and Systematics 32:127-157.
Porter E. E., B. R, Forschner, and R. B. Blair. 2001. Woody vegetation and canopy fragmentation along an urban gradient. Urban Ecosystems 5:131-151.
Reale, J. A. 2002. The nesting success of American robins and Northern Cardinals along an urban gradient: implications for avian communities and an assessment of the effectiveness of artificial nests. Thesis, Miami University, Oxford, Ohio, USA.
Rebele, F. 1994. Urban ecology and special features of urban ecosystems. Global Ecology and Biogeography Letters 4:173-187.
Reitsma, L. R., R. T. Holmes, and T. W. Sherry. 1990. Effects of removal of red squirrels, Tamiasciurus hudsonicus, and eastern chipmunks, Tamias striatus, on nest predation in a northern hardwood forest: an artificial nest experiment. Oikos 57:375-380.
Reynolds, R. T., J. M. Scott, and R. A. Nussbaum. 1980. A variable circular-plot method for estimating bird numbers. Condor 82:309-313.
Ricklefs, R. E. 1969. An analysis of nesting mortality in birds. Smithsonian Contributions to Zoology 9:1-48.
Ricklefs, R. E., and D. Schluter. 1993. Species diversity in ecological communities: historical and geographical perspectives. University of Chicago Press, Chicago, Illinois, USA.
Roper, J. J. 1992. Nest predation experiments with quail eggs: too much to swallow? Oikos 65:528-530.
Rudnicky, J. L., and M. J. McDonnell. 1989. Forty-eight years of canopy change in a hardwood-hemlock forest in New York City. Bulletin of the Torrey Botanical Club 116:52-64.
Shannon, C. E., and W. Weaver. 1949. The mathematical theory of communication. University of Illinois Press, Urbana, Illinois, USA.
Sorace, A. 2002. High density of bird and pest species in urban habitats and the role of predator abundance. Ornis Fennica 79(2):60-71.
Storaas, T. 1988. A comparison of losses in artificial and naturally occurring Capercaillie nests. Journal of Wildlife Management 52:123-126.
Stein B. A., L. Kutner, and J. Adams. 2000. Precious heritage. Oxford University Press, Oxford, UK.
Thorington, K. K., and R. Bowman. 2003. Predation rates on artificial nests increase with human housing density in suburban habitats. Ecography 26(2):188-196.
U.S. Census Bureau. 1990. Land area, population, and density for places: 1990. U.S. Census Bureau, Population Division, Population and Housing Programs Branch, Washington D.C., USA.
U.S. Census Bureau. 2001. Statistical abstracts of the United States. GPO, Washington, D.C., USA.
U.S. Department of Housing and Urban Development. 2000. The state of the cities. HUD, Washington, D.C., USA.
Vale, T. R., and G. R. Vale. 1976. Suburban bird populations in west-central California. Journal of Biogeography 3:157-165.
Vitousek, P. M., H. A. Mooney, J. Lubchenco, and J. M. Melillo. 1997. Human domination of Earth's ecosystems. Science 277:494-499.
Whitney, G. G., and S. D. Adams. 1980. Man as maker of new plant communities. Journal of Applied Ecology 17:341-448.
Address of Correspondent:
Robert Blair
Department of Fisheries, Wildlife, and Conservation Biology
University of Minnesota
St. Paul, Minnesota 55108 USA
Phone: (612) 624-2198
Fax: (612) 625-5299
blairrb@umn.edu
![]()