






The following is the established format for referencing this article:
O'Gorman, C. J., L. P. Bentley, C. McKay, M. Purser, and K. M. Everly. 2022. Examining abiotic and biotic factors influencing specimen black oaks (Quercus kelloggii) in northern California to reimplement traditional ecological knowledge and promote ecosystem resilience post-wildfire. Ecology and Society 27(2):19.ABSTRACT
California black oak, Quercus kelloggii, plays an important role in the lifeways of many Indigenous tribes throughout California. Native peoples tend black oaks using traditional ecological knowledge (TEK) to encourage the development and proliferation of specimen oaks. These mature, large, full crowned trees provide a disproportionate amount of ecosystem services, including acorns and habitat, when compared to smaller black oaks. Altered approaches to land management and the cessation of frequent low intensity cultural burns places these specimen oaks at risk from encroachment, forest densification, and severe wildfire. This project is a collaboration between academic researchers and a Native Advisory Council to examine abiotic and biotic factors influencing Quercus kelloggii to reimplement traditional ecological knowledge and promote ecosystem resilience post-wildfire. Data were collected from 55 specimen black oaks at Pepperwood Preserve in northeast Sonoma County. Specifically, we classified specimen oak growth habitat by measuring specimen oak crown area and live crown ratio, the size and number of surrounding trees, and amount of surface and ladder fuel loads. The preserve burned in both the Tubbs Fire in 2017 and the Kincade Fire in 2019, the latter of which occurred three months after the completion of the initial data collection. Immediately following the Kincade Fire we measured scorch height and related our abiotic and biotic variables to fire severity. Forest densification was found to have a significant negative effect on both canopy area (p = 0.003) and live crown ratio (p = 0.038) of the specimen oaks. Densification did not affect surface and ladder fuel load accumulation since the Tubbs Fire in October 2017 (p > 0.05). Neither surface and ladder fuels nor forest densification variables significantly affected scorch height on the specimen oaks (p > 0.05) following the Kincade Fire.INTRODUCTION
California black oak (Quercus kelloggii) has the widest distribution and altitudinal range of all the oaks in the western U.S., extending over approximately 780 miles from Eugene, Oregon to Baja California (McDonald et al. 1990). Black oak is a keystone species that provides numerous ecosystem services (Crotteau et al. 2015, Long et al. 2017). In California oak woodlands, over 330 species of animals depend on support from oaks for at least one life stage (Barrett 1980).
Black oaks are considered to be a “cultural keystone” species for their role in Indigenous peoples’ diet, ceremonial practices, and daily life (Garibaldi and Turner 2004, Long et al. 2016, 2017). As a food source, black oak acorns are highly desired for their flavor, high oil content, ease to pound and grind, as well as their storability (Lee 1998, Long et al. 2016; M. K. Anderson 1993, unpublished report). The act of gathering acorns is vital for cultural transmission and remains an important social activity for many Tribal members (Anderson 2005, Long et al. 2016). Black oak acorns are incorporated into dances, rituals, and ceremonies and the species appears in Native mythology (Long et al. 2016). Because of their cultural importance, Native peoples throughout present day California use traditional ecological knowledge (TEK) to create ecological conditions that favor black oak survival and persistence while treating the ecosystem as a whole (Anderson 2005, Lake et al. 2017, Long et al. 2017). TEK has been described as “a cumulative body of knowledge, practice, and belief, evolving by adaptive processes and handed down through generations by cultural transmission, about the relationship of living beings (including humans) with one another and with their environment” (Berkes et al. 2000:1252). These cultural practices are much more than land stewardship techniques, but rather, integral pieces of Indigenous culture that reflect a reciprocal relationship with the land.
Despite its vast range, drought adaptability, and cultural significance, black oaks are threatened by altered approaches to land management. Colonization and the forced removal from and cessation of Native engagement with the land resulted in large ecological shifts in California including habitat reduction for many native plants (Kimmerer and Lake 2001, Anderson 2005, Codding and Bird 2013). California Tribal communities have been dispossessed of their traditional lands and their ability to maintain cultural practices and autonomy stifled through subjugation of racist land use laws, as well as outright genocide (Fenelon and Trafzer 2014, Norgaard 2014, Madley 2016). Indigenous peoples’ ability to practice their culture depends on access to the land and the continued resilience of the plants and plant communities on which they have depended for millennia (Berkes et al. 2000, Anderson 2005, Garibaldi and Turner 2004, Long et al. 2016, 2017).
Native peoples create intimate multi-generational relationships with individual specimen black oak trees that include burning, tending, nurturing, and gathering (Anderson 2005). In Sonoma County, California at Pepperwood Preserve, the Native Advisory Council of Pepperwood wishes to return these practices to the land. Pepperwood Preserve is located on the traditional ancestral territory of the Wappo (Milliken 2007). Black oak holds a position of great cultural importance and is the Wappo tribe’s preferred acorn for eating. As such, this project was developed as one of the initial steps in facilitating the reimplementation of these reciprocal stewardship practices. Our research was a collaboration between Pepperwood’s Native Advisory Council, Clint McKay (Wappo/ Pomo/ Wintun), Chair, Lucy McKay (Pomo and Northern Sierra Miwok), Dr. Brenda Flyswithhawks (Tsalági), Tek Tekh Gabaldon (Mishewal Wappo), and L. Frank Manriquez, (Tongva and Ajachemem and Rarámuri), Pepperwood’s management team, and researchers at Sonoma State University.
In meetings with Sonoma State researchers and the Pepperwood management team, Pepperwood’s Native Advisory Council expressed concerns regarding the presence and regeneration of specimen black oaks on the preserve, not just the abundance of the black oak species. Specimen oaks have been defined as large, old, healthy trees with broad low crowns (Long et al. 2017). The Native Advisory Council explained that this growth habit makes it easier to gather acorns and also supports other culturally significant understory plants. These trees provide a disproportionate number of ecocultural services when compared to smaller black oaks (Long et al. 2017). Clint McKay explained the following:
[A] specimen oak does not come with age. It comes with the overall health of the tree and the tree’s ability to take its rightful place within our natural world in the spirit of reciprocity. The specimen oak benefits from its microenvironment and in return can reciprocate its bounty to the natural world; humans, animals and other plant communities. Overcrowding or densification causes the trees to not reach their full potential. We get small, spindly trees that lack strength and the ability to produce the quality and quantity of acorns needed to support other life forms.
Like numerous Indigenous groups around the world, Sonoma County Tribal communities have difficulty accessing the plants necessary for cultural practices, such as basket weaving and maintaining a traditional diet, because of public land gathering prohibitions, the cessation of cultural burns, limited access to land, and habitat destruction (Garibaldi and Turner 2004, Anderson 2005, Shebitz et al. 2009, Long et al. 2017). A very limited amount of land in Sonoma County is under Tribal jurisdiction. As a result, Tribal communities cannot possibly sustain themselves on such limited land resources alone. Traditional gathering places beyond Tribal lands are not always accessible, which further restricts the traditional foods and cultural resources contained therein. Lack of access to traditional foods and the use of pesticides and herbicides on adjacent agricultural lands directly affect both overall Tribal health and the conditions of the few gathering places that remain open to Tribal peoples. Policies, such as gathering restrictions on abalone and salmon, also affect Native peoples (Clint McKay, personal communication). Despite all the challenges, Tribal communities continue to practice their cultural traditions and pass knowledge through the generations. This cultural resilience is expressed in many ways including efforts to revitalize language and traditional crafts like basket weaving. It is also expressed by connecting traditional stewardship practices with modern day political movements like climate activism. A key component to cultural resilience is working to restore both access to the land and the ability to engage in traditional cultural practices.
Present day conservation depends on understanding the ecological and cultural history of a place, including cultural practices and the roles and benefits they played in a given ecosystem (Kimmerer and Lake 2001, Keeley 2002, Hayashida 2005, Codding and Bird 2013). Historically, researchers rooted in the binary anthropological distinction between hunter gatherers and agricultural societies missed Indigenous peoples’ influence across many habitats of the California landscape and were unaware of the importance of active management in California’s ecosystems (Anderson 2005, Shebitz et al. 2009, Codding and Bird 2013, Lepofsky et. al 2017). Indigenous people act as keystone species in numerous ecosystems around the world through the use of fire (Kimmerer and Lake 2001, Miller and Davidson-Hunt 2010, Codding and Bird 2013, Christianson 2014). Fire has numerous beneficial effects on a landscape scale, including plant germination, plant health, and maintaining habitat heterogeneity (Kimmerer and Lake 2001, Keeley 2002, Anderson 2005, Abrams and Nowacki 2008, Codding and Bird 2013, Kelly and Brotons 2017, Anderson and Keeley 2018). The use of fire to increase abundance and production of masting tree species has been documented across North America (Abrams and Nowacki 2008). In California, intensification of the use of oak and material correlates that accompany acorn processing such as bedrock mortars, pestles, and acorn macrofossils date from ~6500 to 4300 cal BC (Hilderbrant 2007). Evidence from paleoecology shows frequent cultural burns also occurred across much of California during the mid to late Holocene, which is supported by the ethnographic and archaeological record (Keeley 2002, Anderson 2005, Stephens et al. 2007, Lightfoot et al. 2013, Christianson 2014, Lightfoot and Cuthrell 2015, Anderson and Keeley 2018). Burning and similar cultural practices have significant time depth and have been a component of the black oaks’ environment for millennia.
Burning practices ceased in Sonoma County within the last 200 years after the imposition of fire suppression policies (Barnhart et al. 1996, Hastings et al. 1997, Minnich et al. 2000, Kimmerer and Lake 2001). Density of oak stands has increased since the time of Euro-American settlement (1850–1910), and the state’s adoption of a fire suppression model has increased oak mortality (Barnhart et al. 1996, Hunter and Barbour 2001, Schriver et al. 2018). Fire suppression has allowed for forest densification by Douglas-fir (Pseudotsuga menziesii) and hardwood species, which increases drought sensitivity in oaks (Barnhart et al. 1996, Hunter and Barbour 2001, Cocking et al. 2012, Long et al. 2018, Gedalof and Franks 2019). Competition by neighboring trees, including hardwood species, results in loss of acorn production and higher rates of mortality (Asbjørnsen et al. 2007, Marcos et al. 2007, Oheimb et al. 2011, Long et al. 2018). In addition, forest densification increases the chance of oak mortality due to shade induced stress and an increase in ladder fuels (Barnhart et. al. 1996, Cocking et al. 2012). Increased ladder fuels enable fire to climb into oak canopies thereby increasing the chance of top kill and mortality (Horney et al. 2002). The absence of fire allows for fuel load accumulation that can result in higher fire severity thereby increasing the chance of top kill in a fire event, and unintended injury to mature oaks during prescribed fires (Long et al. 2018, Nemens et al. 2019; Fig. 1).
Black oaks rely on frequent, low intensity fires and this fire regime supports the development and proliferation of old, large black oaks with broad, low crowns (Skinner et al. 2006, Cocking et al. 2012, Long et al. 2016, Hammett et al. 2017). The Native Advisory Council and other cultural practitioners within California have expressed the cultural and ecological importance of these specimen oaks and the ecocultural services they provide (Cocking et al. 2012, Long et al. 2016, 2017, Crotteau et al. 2015). Fire suppression and its effects, including forest densification, place existing specimen oaks at risk and discourage the regeneration and development of future generations of legacy trees (Cocking et al. 2012, Long et al. 2016, 2017, Nemens et al. 2018).
METHODS
Objectives of the Native Advisory Council and research design
The Native Advisory Council asked that Sonoma State researchers identify specimen black oaks at Pepperwood Preserve and record the ecological conditions of the trees. Based on the concerns expressed by the Native Advisory Council, a research design was created to examine the effects of forest densification on specimen oak crown shape. The accumulation of fire fuels since the Tubbs Fire in 2017 was also measured. Scorch height on the specimen trees was measured after the Kincade Fire, which occurred during the field research in 2019, to determine the impact of our measured biotic factors on fire severity around these specimen oaks.
Study site
Pepperwood Preserve consists of 1263 hectares (3120 acres) located northeast of the city of Santa Rosa in the Southern Mayacamas Mountains (Gillogly et al. 2017). Elevation on the Preserve ranges from 56 to 475 meters (185–1560 feet). Though situated 40 kilometers (25 miles) east of the Pacific Ocean, maritime influences such as coastal fog impact the preserve (Gillogly et al. 2017). In total, 543 native species have been collected and documented on the Preserve, 18 of which are endemic (DeNevers 2013, Halbur et al. 2013, Gillogly et al. 2017). Oak woodland stretches across 394 hectares (974 acres) of the Preserve making it the dominant vegetation type. The main overstory oak species are live oak (Quercus agrifolia var. agrifolia), blue oak (Quercus douglasii), black oak, Oregon white oak (Quercus garryana), and valley oak (Quercus lobata; Gillogly et al. 2017). Other associated overstory species include Douglas-fir, tanoak (Notholithocarpus densiflorus), madrone (Arbutus menziesii), and bay (Umbellularia californica).
Location and determination of specimen oaks
To determine the location of specimen oaks within the Preserve, cruising surveys were performed by vehicle and foot along main roads. LiDAR and hyperspectral derived vegetation maps were also used to identify areas of high black oak abundance (Fig. 2). Initially, the search covered a 100 meter swath on either side of major roads and trails to ensure easy access by cultural practitioners, but eventually extended to more remote locations after mapping the specimen black oaks adjacent to roads and trails. Once a potential specimen oak was found, its geospatial location was recorded (Garmin GPS), the tree was photographed and marked by placing a tagged rebar stake into the ground two meters from the north side of the trunk.
All 55 specimen oaks identified were within the Tubbs Fire perimeter which burned roughly 95% of Pepperwood Preserve in October 2017. The Tubbs Fire perimeter encompassed roughly 14,895 ha, and more than 25% burned in both medium and high severity (Ackerly et al. 2019). After the initial data collection was completed, roughly 60% of the Preserve burned in the Kincade Fire in October 2019. Of the 55 specimen oaks, 37 were within the Kincade Fire perimeter and two fell during the fire event. (Fig. 2)
Ecological field measurements
Tree height of each specimen oak was measured using an inclinometer. Crown height was determined by measuring from the base of leafy canopy to top of canopy. The two crown dimension variables measured were crown area and live crown ratio. To determine crown area, the major and minor axes of the crown were measured with a meter tape on the ground underneath the crown. These measurements were then used to solve for the area of an ellipse. Live crown ratio was determined by dividing the total tree height by the canopy base height.
Diameter at breast height (DBH) was recorded for each tree. DBH was measured 1.3 meters from the base of the tree on the uphill side, perpendicular to the trunk axis and to the nearest tenth of a centimeter. To determine fire risk to each specimen oak, surface fuels and ladder fuels were measured around each tree. Three randomly selected and equidistant Brown’s transects originating from the base of the specimen oak were used to quantify surface fuels. Ladder fuels were quantified by photographing a 4 m tall banner located 15 meters from the base of the tree at the end of each Brown’s transect and then quantifying the area covered by both dead and live branches using the program ImageJ (Kramer et al. 2016). Any branches belonging to the specimen oak were not included in the analysis.
To quantify forest densification, all existing and potential forest canopy trees larger than two meters in height were surveyed in a 15 meter circular plot with the specimen oak at the origin. Species, DBH, tree condition (live, dead, or top kill), and distance to specimen oak were recorded. Canopy position was determined for these trees (by the same individual observer throughout the experiment). Canopy positions were categorized as understory (UN) if less than 1/3 of the crown height penetrates the bottom of the specimen oak canopy, emergent (EM) if 2/3 of the crown height is within the canopy of the oak, and oak overstory (O) if 1/3 of the crown height is level with the top of the oak canopy (Cocking et al. 2012). Within two weeks following the Kincade fire, scorch height was measured on all specimen oaks using an inclinometer or meter tape when possible.
Statistical analyses
A hierarchical set of models was fit to analyze the effect of forest densification on crown area and live crown ratio. The initial model used the following forest densification variables as factors: total DBH of surrounding trees and the average distance of surrounding trees to the specimen oak tree. Average distance to the specimen oak was not found to be significant (p > 0.05) in any of the models and, as such, was excluded from subsequent analyses. When the total DBH of surrounding trees was significant, a new model was used to disentangle the effects of its two component parts: mean DBH of surrounding trees (e.g., area) and number of surrounding trees (e.g., abundance). If either mean DBH of surrounding trees or number of surrounding trees was significant, a final model was run where each variable was then categorized by canopy positions (overstory, emerging, and understory) to determine if the canopy position of the surrounding trees affected live crown ratio and crown area.
Two separate generalized linear models (GLM) were used to determine the effect of forest densification on surface fuels and ladder fuels. One hour, 10 hour, and 100 hour surface fuels were included in the analyses. Forest densification variables included total DBH of surrounding trees, number of surrounding trees, and average distance of surrounding trees to the specimen oak.
To determine the effect of the Kincade Fire on each specimen oak tree, scorch height was measured as the height from the base of the specimen oak tree to the highest visible char mark. It was then standardized by dividing scorch height by total tree height. The effects of forest densification variables and fire fuels (tons per acre of fine surface fuels and total amount of ladder fuels) on scorch height were evaluated in separate models. All analyses were run in JMP 14.
RESULTS
Plot composition and structure
In total, 428 canopy trees from 8 species were present in the 55 plots. Mean DBH of specimen oaks was 100.37 cm (range = 65.3 cm to 168.6 cm). The mean height of specimen oaks was 14.82 m (range = 10.28 m to 23.33 m). Sixty-nine percent of all surrounding tree species were oaks (Quercus spp.). Coast live oak was the most abundant species (27% of surrounding trees; 116 trees; present in 65.45% of plots; mean DBH of 54.26 cm). The other tree species (listed in order of abundance) were black oak (23% of surrounding trees; 100 trees; present in 63.64% of plots; mean DBH of 55.04 cm), madrone (15% of surrounding trees; 65 trees; present in 40% of plots; mean DBH of 12.96 cm), Oregon white oak (11% of surrounding trees; 48 trees; present in 36.36% of plots; mean DBH of 43.17 cm), bay (8% of surrounding trees; 34 trees; present in 36.36% of plots; mean DBH of 18.45), Douglas-fir (7% of surrounding trees; 30 trees; present in 32.73% of plots; mean DBH of 22.22 cm), blue oak (6% of surrounding trees; 25 trees; present in 16.36% of plots; mean DBH of 48.73), and tanoak (0.23% of surrounding trees; 1 tree; present in 1.81% of plots; DBH of 13 cm).
All canopy trees were categorized by canopy position. There were 168 understory trees (mean DBH = 15.12 cm), 80 emerging trees (mean DBH = 50.05 cm), and 178 overstory trees (mean DBH = 61.23 cm). The species accounting for the largest percentage of each canopy position were understory (35.19% madrone), emerging (36.71% coast live oak), and overstory (34.83% black oak).
Surface, ladder fuels, and scorch height
Across all plots, mean surface fuels were 5.4 tonnes per hectare (range = 2.7 to 8.5 tonnes per hectare). Ladder fuels (dead or live vegetation from 0 to 4 m in height) ranged from 0% to 48% across all plots. Mean scorch height was 4.5 m (range = 0.3 m to 15.8 m). Scorch height was made equivalent to total tree height for the two specimen oaks that fell during the Kincade fire event. None of the forest densification variables had a significant effect on the amount of fine surface fuels per square hectare or the amount of ladder fuels pre-Kincade Fire (p > 0.05). The forest densification variables, fine surface fuels, and ladder fuels did not have a significant effect on scorch height post-Kincade Fire (p > 0.05).
Forest densification on crown area
Our initial model showed a strong negative effect of total DBH of surrounding trees on specimen oak crown area (F1, 50 = 9.98, p = 0.0027; Fig. 3). In the subsequent model, both mean DBH of surrounding trees (F1, 51 = 7.33, p = 0.0092), the number of surrounding trees (F1, 51 = 7.58, p = 0.0082), and their interaction (F1, 51 = 6.27, p = 0.016) had a negative effect on specimen oak crown area (Fig. 3). The final model found that the number of emerging trees had a marginally significant negative effect on crown area (F1, 20 = 4.17, p = 0.055). Although we detected a marginally significant negative effect of the number of emerging trees, we note that for 37 of the specimen trees there were no surrounding emerging trees.
Forest densification on live crown ratio
The initial model showed that total DBH of surrounding trees negatively affected specimen oak live crown ratio (F 1, 50 = 4.55, p = 0.0378). The subsequent model showed a marginally significant negative effect of number of surrounding trees on specimen oak live crown ratio (F1, 51 = 3.73, p = 0.0591). The final model, including canopy positions, indicated that the number of overstory trees had the greatest negative effect on specimen oak live crown ratio (F1, 20 = 8.37, p = 0.009).
DISCUSSION
Overall goal of tending trees and incorporating TEK specifically at Pepperwood
In this collaborative project, we sought to find and map specimen black oaks for the reimplementation of TEK within Pepperwood Preserve. Based on expressed concerns of the Native Advisory Council regarding the regeneration and crown shape of legacy trees, we measured ecological variables to identify abiotic and biotic factors influencing specimen oak growth and habitat post-wildfire. Protecting existing legacy trees and restoring black oak ecosystems at Pepperwood Preserve presents the opportunity to partner with local Tribal peoples and increase their access to land while promoting ecosystem restoration (Long et al. 2017, 2020).
Understanding the effects of wildfire on tree structure
Our study site experienced two sequential wildfires (Tubbs and Kincade) over a short temporal scale. Both fires involved wind speeds of over 95 kilometers per hour. Such fires spread quickly and are unpredictable because of erratic movements (Werth et al. 2011). This “extreme fire behavior” is affected by numerous unseen factors such as wind shear and atmospheric stability (Werth et al. 2011). The lack of a significant effect of the forest densification variables on scorch height suggests that fire severity may be operating on a larger spatial scale than the individual surrounding tree variables and that there is more complexity to fire severity aside from the immediate environment of the tree. Larger spatial scale analysis may provide insights to fire movements and risk assessment in such extreme conditions as well as the ways that larger landscape level ecological variables such as tree density affect fire severity and tree mortality (Eugenio et al. 2016). In less extreme conditions, the immediate environment around a focal tree would have been more significant. Fuel loading was low and relatively homogeneous throughout the 55 plots, which we attributed to the first wildfire in 2017 (Tubbs Fire). Fire fuel treatments have been shown to reduce the severity of other fire events and reduce the chance of canopy fire (Stephens et al. 2009). We note that many of the specimen oaks were in relatively good condition prior to the fires and that the surrounding environments were not overly impacted by densification. This may have affected the lack of significant relationship between densification and scorch height. The study was limited to black oaks within the Tubbs Fire perimeter, therefore the analysis describes the effects of a reburn rather than a general wildfire. These constraints on the analysis may understate the importance of pre-wildfire treatments on desired outcomes.
This short fire return interval coupled with the small spatial scale of our study presents a challenge when looking at fuel load accumulation over time and fire movement, but also a unique opportunity to study the effect of repeated wildfires on black oak regeneration and species composition at the preserve.
High severity fire favors black oak over fire sensitive conifers and leads to increased black oak composition in the overstory, but can result in top kill of specimen oaks and the loss of the cultural and ecological services they provide (Cocking et al. 2014, Hammett et al. 2017, Nemens et al. 2018). A separate study at Pepperwood Preserve found that black oaks that burned during the Tubbs Fire experienced high levels of mortality relative to other hardwood species, and that basal resprouting was common (Ackerly et al. 2019). The high severity Tubbs and Kincade fires can be viewed as a “reset” and present an opportunity to follow up with increased stewardship (Long et al. 2016). Frequent fire combined with grazing may reduce or eliminate the shrub layer resulting in an oak savanna (George and Alonso 2008).
Understanding the effects of forest densification on tree and stand structure at Pepperwood
Surrounding overstory trees accounted for the greatest negative effect on live crown ratio of specimen oaks and larger tree size magnified the negative effect on black oak crown diameter. Other studies have also found that competition for light, space, and water represses crown growth (Harrington and Devine 2006). Encroachment by conifers and other hardwood species greatly impacts tree growth and vitality, especially in shade intolerant species such as black oak (McDonald 1969, McDonald et al. 1990, Cocking et al. 2012, Schriver et al. 2018). Acorn yield increases with crown diameter in California black oak (McDonald et al. 1990) and other oak species (Rose et al. 2012) and therefore repressed crown growth can equate to a reduction in acorn production, thereby negatively affecting the trees and the ecological and cultural services they provide.
Vegetation dynamics of black oak woodlands at Pepperwood and the surrounding area are less studied than those in the northern part of the state and southern Oregon, where conifers are generally dominant in the overstory (George and Alonso 2008, Cocking et al. 2012, 2015). In both locations, oaks serve as nurse trees and increased canopy density of oak woodlands assists in recruitment of Douglas-firs and other later successional canopy trees (McDonald 1969, McDonald et al. 1990, Barnhart et al. 1996, Spector and Putz 2006). Encroachment, especially the piercing of oak crowns by Douglas-firs, results in loss of tree vigor, decreased resilience in fire events, crown dieback, and potentially senescence of the focal oak (Barnhart et al. 1996, Spector and Putz 2006, Devine et al. 2007).
A closed oak canopy supports an increasingly mesic understory, which facilitates the replacement of black oak by species such as Douglas-fir and bay, thereby creating a mixed evergreen forest (Wainwright and Barbour 1984, Barnhart et al. 1987, Hastings et al. 1997). A study conducted in Santa Rosa observed black oak woodland in what may represent several successional phases ranging from open oak woodland, an increase of bay and Douglas-fir in the understory, and finally bay and Douglas-fir dominated canopy (Wainwright and Barbour 1984). Rapid Douglas-fir expansion and increased canopy density has occurred in Sonoma County in the past 100 years because of fire suppression and grazing cessation (Barnhart et al. 1996, Hastings et al. 1997). Madrone, juvenile Douglas-firs, and bays accounted for the majority of understory trees in our plots. Shared mycorrhizal associates between madrone and Douglas-fir may contribute to recruitment of the conifer where madrone is present, while increased bay facilitates the spread of the Sudden Oak Death pathogen (Phytophthora ramorum); both of these dynamics will place additional stress on the specimen black oaks (Hunter and Barbour 2001, Meentemeyer et al. 2004). Though grazing and mechanical Douglas-fir removal treatments occur on the preserve, increased canopy density and grassland reduction has been observed over the past 70 years (A. Dawson 2008, unpublished report). This combined with an increase of madrone, bay, and Douglas-fir in the understory suggest gradual composition shifts from fire-tolerant and shade intolerant species, such as black oak and other hardwoods, to less fire-tolerant more shade tolerant hardwoods, and the nursing of young Douglas-fir trees. Many fire dependent plant communities across North America have undergone a similar process, referred to as mesophication. Mesophication can result in fire regime change and alter fire behavior and effects by creating damper, more shaded understory conditions with less flammable fuelbeds (Nowacki and Abrams 2008, Engber and Varner 2012). Of all the California oaks, black oak leaf litter is among the most flammable and generates the deepest fuelbeds (Engber and Varner 2012). Burning of black oak litter was found to result in heavy fuel consumption, brief flaming and smoldering, and low intensity fire (Engber and Varner 2012). The majority of encroaching overstory and emerging trees were live oaks, which were found to repress fire (Engber and Varner 2012). It is possible that this current trajectory will continue without the implementation of additional stewardship practices.
Recommendations for stewardship in light of TEK
The Pepperwood Native Advisory Council advocates for increased prescribed burning and thinning as a means to prepare the land for cultural burning. An important difference between prescribed and cultural fire is that prescribed burns primarily focus on creating fire breaks and reducing fuels, whereas a cultural burn takes an ecosystem level approach to promote the health of the natural environment as a whole, including plant, animal, and human life. A unique aspect about the ongoing project at Pepperwood Preserve is that the Native Advisory Council and Pepperwood staff collaborate on the entire restoration process. (Clint McKay, personal communication).
This twofold approach to restoring the land and protecting specimen oaks begins with fuel reduction in the form of mechanical removal, pile burning, and prescribed fire, followed by the continued use of fire to stave off further degradation and support ecosystem resilience. In addition to preparing the land for cultural burns, our results suggest that mechanical removal of overstory and emerging trees encroaching on specimen oaks may be the best first step to encourage increased crown area and live crown ratio, both of which are desired by today’s traditional gatherers including those represented on the Pepperwood Native Advisory Council (Cocking et al. 2012, Crotteau et al. 2015, Long et al. 2016, 2017; Clint McKay, personal communication). Thinning in oak woodlands has been shown to reduce water stress (Moreno and Cubera 2008) and release from encroaching trees increases epicormic branching in oaks and overall growth, thereby increasing crown dimensions (McDonald 1969, Harrington and Devine 2006, McDonald and Vaughn 2007) and mast products in black oak and other oak species (Healy 1997, Healy et al. 1999). Mechanical removal followed by controlled or pile burning is an effective fuel reduction treatment (Hastings et al. 1997, Stephens et al. 2009). Because the majority of observed Douglas-firs were seedlings and juveniles (mean DBH 20 cm), controlled burns would be an effective tool in mitigating conifer encroachment (Hunter and Barbour 2001, Cocking et al. 2012), as well as continuing the efforts of mechanical removal.
Following mechanical removal of fuels and encroaching trees, and then prescribed fire, cultural burns similar to the frequent, low severity fire regimes seen in the ethnographic and archeological record (Hastings et al. 1997, Keeley 2002, Anderson 2005, Christianson 2014, Lightfoot and Cuthrell 2015) would protect specimen black oaks and facilitate the development of mature black oak stands in part by discouraging conifer recruitment and hardwood densification after severe fire events (Cocking et al. 2012, Crotteau et al. 2013, Long et al. 2016, 2017, Hammett et al. 2017, Nemens et al. 2018). In the context of TEK, cultural burning fits in a larger system of ecological management and Tribal stewardship strategies. For example, the use of fire during certain times of the year may also promote native, culturally significant understory plants and help control pests and pathogens while simultaneously restoring Indigenous land relationships (Kimmerer and Lake 2001, Huffman 2013, Long et al. 2016, Lake et al. 2017, Anderson and Keeley 2018). In the fall of 2020, the Native Advisory Council and the Pepperwood management team began mechanical removal followed by pile burning on the preserve, with the intention of reimplementing cultural fire when the land is ready.
Many researchers encourage us to view TEK not as a static system locked in the past but as a dynamic system rooted in adaptation (Pearce et al. 2015). Pearce and colleagues’ work (2015) views TEK as an element of adaptive capacity or resilience and shows how a peoples’ ability to adapt may depend on their ability to continue to develop TEK in the context of a changing environment. It is essential to remember that major biotic and abiotic changes have occurred since precontact times, including the introduction of invasive plants, increased fuel accumulation, and stand density, and a warming climate. Western science and TEK can work cooperatively to assess California ecosystems in their current state and create unique stewardship plans for the particular needs of a given site that account for ongoing climatic and ecological change.
A way forward
To foster resilient ecosystems that support Tribal values a multifaceted and diverse approach that incorporates Tribal communities, land management agencies and organizations, government, and the public is needed to create unique plans for different social and ecological conditions throughout the state (Lake et al. 2017, Long et al. 2017, Long and Lake 2018). Government action and policy, at the federal, state, and local levels, impact Tribal engagement with and access to the land. For example, the Northwest Forest Plan (NWFP) helped to facilitate increased collaboration with Tribal communities whose lands are within the designated boundary (Long and Lake 2018). Though the plan did not adequately include Tribal stewardship of ecocultural resources, subsequent land management planning rules prioritize the protection and management of sites and resources of cultural importance (Long and Lake 2018). Obstacles to Tribal stewardship on the land take many forms and range from the federal level down to local restrictions on burning such as air-quality regulations (Quinn-Davidson and Varner 2012, Long et al. 2017, Long and Lake 2018). Despite this, other successful partnerships focused on TEK and black oak restoration have formed throughout the state including work by the Honorable Ron Goode and the North Fork Mono Tribe with the Sierra National Forest and Greenville Rancheria in collaboration with Plumas National Forest (Long et al. 2017). These partnerships and others exemplify the dynamic and inclusive active stewardship strategies needed to cultivate social and ecological resilience and simultaneously promote ecocultural resources.
It is essential that the agencies, organizations, and other entities with power and funding reach out in a culturally competent and sensitive manner to include Tribal communities in decision making and fund local Tribal action on the land. Understanding the importance of place and cultural competency around Indigenous worldview will aid in collaborations with Tribal peoples, and incorporating local Tribal peoples into leadership roles will aid in the decolonization of land stewardship (Muller 2003, Dockry et al. 2018). Collaboration between Tribal communities and land stewardship organizations is mutually beneficial. Acknowledging Tribal authority and legitimizing TEK has political benefits for Tribal groups, and Tribal involvement will benefit ecosystems and society by incorporating the wealth of knowledge embedded within TEK into land stewardship programs. Successful partnerships have formed between land management agencies and indigenous peoples in Australia, Canada, and California, many of which involve TEK of fire (Muller 2003, Shebitz et al. 2009, Christianson 2014, Robinson et al. 2016, Lake et al. 2017).
In conclusion, collaborating with and providing the opportunity for Tribal communities to practice TEK will increase access to land and biodiversity, keep California’s diverse vegetation types from undergoing successional change, and promote the growth and resilience of culturally significant plants (Anderson 2005, Burr 2013, Fenelon and Trafzer 2014, Norgaard 2014, Armstrong and Veteto 2015, Livingston et al. 2016). This has far reaching implications on Tribal peoples’ well-being, their ability to engage in cultural practices, and ecosystem health, because many cultural practices are inextricably linked to California’s biodiversity and aid in its preservation. Active stewardship of California ecosystems and incorporating Tribal communities in land stewardship action and decision making can benefit the health and well-being of local tribes, the larger surrounding communities, and ecosystems alike.
Clint McKay shared his reflections on the importance of this work:
Native people talk about seven generations. We live our life today based on the effects our actions will have seven generations down the line. After the Tubbs and Kincade fire I got to thinking about the stewardship of this land. I started counting back from my grandchildren, and if you go back seven generations that was the last time my people were able to steward this land according to our cultural values and traditions. To us these big wildfires are like a reawakening of our culture and traditions and it’s given this western world an opportunity to push the reset button and start over. For seven generations my people have been denied the opportunity to steward this land according to our culture and traditions. Unfortunately, it took these devastating wildfires but now our scientific methods are starting to be heeded. People are starting to ask for it and respect it. It is an opportunity for learning, sharing, and bringing us back to a place of center not for pointing fingers. We get another opportunity; what are we going to do with it now?
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.AUTHOR CONTRIBUTIONS
CJO: Conceptualization, Methodology, Formal Analysis, Investigation, Data Curation, Writing: Original Draft, Visualization, Project Administration, Funding Acquisition
MP: Writing: Review and Editing
CM: Conceptualization, Methodology, Writing: Interviews and Review
KME: Investigation, Data Curation
LPB: Conceptualization, Methodology, Resources, Writing: Review and Editing, Visualization, Supervision, Project Administration, Funding Acquisition
ACKNOWLEDGMENTS
Funding for this work was awarded to Cory O'Gorman from Northern CA Botanists CNPS Milo Baker Chapter, Sonoma State University’s Center for Environmental Inquiry Norwick Fund and RSCA Program. Daniel Crocker was instrumental in assisting with statistical analyses and we are thankful for his participation in this project and suggestions to improve the manuscript. We are grateful to Matthew Clark for providing LiDAR and hyperspectral derived vegetation maps to identify areas of high black oak abundance at Pepperwood. We appreciate the Native Advisory Council of Pepperwood, Clint and Lucy McKay, Dr. Brenda Flyswithhawks, Tek Tekh Gabaldon, and L. Frank Manriquez for welcoming us into this project and at their table. We thank Pepperwood Preserve staff for their continued support throughout the project and to Ashley Whipple for assisting with data accessibility. We appreciate comments by Frank K. Lake and an anonymous reviewer whose suggestions greatly improved an earlier versions of the manuscript. We are grateful to the Honorable Ron Goode and Dirk Charley for sharing their expertise and cultural wisdom.
DATA AVAILABILITY
The data that support the findings of this study are openly available in the EDI Data Portal at https://doi.org/10.6073/pasta/6ae5b7174cafbdc95a5b0478081fa11e.
LITERATURE CITED
Abrams, M. D., and G. J. Nowacki. 2008. Native Americans as active and passive promoters of mast and fruit trees in the eastern USA. Holocene 18(7):1123-1137. https://doi.org/10.1177/0959683608095581
Ackerly, D. D., M. Kozanitas, P. Papper, M. Oldfather, and M. Clark. 2019. Mortality and resprouting in California Oak Woodlands following mixed-severity wildfire. International Oaks 30:23-30.
Anderson, M. K. 2005. Tending the wild: Native American knowledge and the management of California’s natural resources. University of California Press, Berkeley, California, USA. https://doi.org/10.1525/9780520933101
Anderson, M. K., and J. E. Keeley. 2018. Native peoples’ relationship to the California chaparral. Pages 79-121 in E. C. Underwood, H. D. Safford, N. A. Molinari, and J. E. Keeley, editors. Valuing chaparral: ecological, socio-economic, and management perspectives. Springer, Cham, Switzerland. https://doi.org/10.1007/978-3-319-68303-4_4
Armstrong, C. G., and J. R. Veteto. 2015. Historical ecology and ethnobiology: applied research for environmental conservation and social justice. Ethnobiology Letters 6(1):5-7. https://doi.org/10.14237/ebl.6.1.2015.313
Asbjørnsen, H., L. A. Brudvig, and M. Tomer. 2007. Ecohydrological implications of removing encroaching woody vegetation from a bur oak savanna. Ecological Restoration 25:58-60.
Barnhart, S. J., R. J. McBride, C. Cicero, P. da Silva, and P. Warner. 1987. Vegetation dynamics of the northern oak woodlands. Pages 53-58 in Multiple-use management of California’s hardwood resources. Pacific Southwest Forest and Range Experiment Station, Berkeley, California, USA.
Barnhart, S. J., J. R. McBride, and P. Warner. 1996. Invasion of northern oak woodlands by Pseudotsuga menziesii (Mirb.) Franco in the Sonoma Mountains of California. Madroño 43(1):28-45.
Barrett, R. H. 1980. Mammals of California oak habitats: management implications. Pages 275-291 in Ecology, management, and utilization of California oaks. Pacific Southwest Forest and Range Experiment Station, Berkeley, California, USA.
Berkes, F., J. Colding, and C. Folke. 2000. Rediscovery of traditional ecological knowledge as adaptive management. Ecological Applications 10(5):1251-1262. https://doi.org/10.1890/1051-0761(2000)010[1251:ROTEKA]2.0.CO;2
Burr, J. L. 2013. Burning across boundaries: comparing effective strategies for collaboration between fire management agencies and indigenous communities in the United States and Australia. Interdisciplinary Studies in the Humanities 5(2012):1-6.
Christianson, A. 2014. Social science research on Indigenous wildfire management in the 21st century and future research needs. International Journal of Wildland Fire 24(2):190-200. https://doi.org/10.1071/WF13048
Cocking, M. I., J. M. Varner, and E. A. Engber. 2015. Conifer encroachment in California oak woodlands. Pages 505-514 in Proceedings of the Seventh California Oak Symposium: managing oak woodlands in a dynamic world. General Technical Report PSW-GTR-251. U.S. Forest Service, Pacific Southwest Research Station, Berkeley, California, USA.
Cocking, M. I., J. M. Varner, and E. E. Knapp. 2014. Long-term effects of fire severity on oak–conifer dynamics in the southern Cascades. Ecological Applications 24(1):94-107. https://doi.org/10.1890/13-0473.1
Cocking, M. I., J. M. Varner, and R. L. Sheriff. 2012. California black oak responses to fire severity and native conifer encroachment in the Klamath Mountains. Forest Ecology and Management 270:25-34. https://doi.org/10.1016/j.foreco.2011.12.039
Codding, B. F., and D. W. Bird. 2013. Forward: a global perspective on traditional burning in California. California Archaeology 5(2):199-208. https://doi.org/10.1179/1947461X13Z.00000000010
Crotteau, J. S., M. W. Ritchie, J. M. Varner, and J. P. Berrill. 2015. Quercus Kelloggii (Newb.) sprout response to fire severity in northern California. Pages 377-386 in Proceedings of the Seventh California Oak Symposium: managing oak woodlands in a dynamic world. General Technical Report PSW-GTR-251. U.S. Forest Service, Pacific Southwest Research Station, Berkeley, California, USA.
Crotteau, J. S., J. M. Varner III, and M. W. Ritchie. 2013. Post-fire regeneration across a fire severity gradient in the southern Cascades. Forest Ecology and Management 287:103-112. https://doi.org/10.1016/j.foreco.2012.09.022
DeNevers, G. 2013. Pepperwood Preserve Vascular Flora. P. Rockwood, M. M. Halbur, and M. Gillogly, editors. Third edition. Dwight Center for Conservation Science at Pepperwood, Santa Rosa, California, USA.
Devine, W. D., C. A. Harrington, and D. H. Peter. 2007. Oak woodland restoration: understory response to removal of encroaching conifers. Ecological Restoration 25(4):247-255. https://doi.org/10.3368/er.25.4.247
Dockry, M. J., S. A. Gutterman, and M. A. Davenport. 2018. Building bridges: perspectives on partnership and collaboration from the U.S. Forest Service Tribal Relations Program. Journal of Forestry 116(2):123-132. https://doi.org/10.5849/jof-2016-106
Engber, E., and J. M. Varner III. 2012. Patterns of flammability of the California oaks: the role of leaf traits. Canadian Journal of Forest Research 42(11):1965-1975. https://doi.org/10.1139/x2012-138
Eugenio, F. C., A. R. dos Santos, N. C. Fiedler, G. A. Ribeiro, A. G. da Silva, Á. B. dos Santos, G. G. Paneto, and V. R. Schettino. 2016. Applying GIS to develop a model for forest fire risk: a case study in Espírito Santo, Brazil. Journal of Environmental Management 173:65-71. https://doi.org/10.1016/j.jenvman.2016.02.021
Fenelon, J. V., and C. E. Trafzer. 2014. From colonialism to denial of California genocide to misrepresentations: special issue on indigenous struggles in the Americas. American Behavioral Scientist 58(1):3-29. https://doi.org/10.1177/0002764213495045
Garibaldi, A., and N. Turner. 2004. Cultural keystone species: implications for ecological conservation and restoration. Ecology and Society 9(3):1. https://doi.org/10.5751/ES-00669-090301
Gedalof, Z., and J. A. Franks. 2019. Stand structure and composition affect the drought sensitivity of Oregon white oak (Quercus Garryana Douglas Ex Hook.) and Douglas-Fir (Pseudotsuga Menziesii (Mirb.) Franco). Forests 10(5):381. https://doi.org/10.3390/f10050381
George, M. R., and M. F. Alonso. 2008. Oak woodland vegetation dynamics: a state and transition approach. Pages 93-104 in Proceedings of the sixth California oak symposium: today’s challenges, tomorrow’s opportunities. General Technical Report PSW-GTR-217. U.S. Forest Service, Pacific Southwest Research Station, Albany, California, USA.
Gillogly, M., C. Dodge, M. Halbur, L. Micheli, C. McKay, N. Heller, and B. Benson. 2017. Adaptive management plan for Pepperwood Preserve: terrestrial biodiversity climate change collaborative. Dwight Center for Conservation Science at Pepperwood, Santa Rosa, California, USA.
Halbur, M., M. Kennedy, D. Ackerly, L. Micheli, and J. Thorne. 2013. Creating a detailed vegetation map for Pepperwood Preserve: terrestrial biodiversity climate change collaborative. Dwight Center for Conservation Science at Pepperwood, Santa Rosa, California, USA.
Hammett, E. J., M. W. Ritchie, and J. P. Berrill. 2017. Resilience of California black oak experiencing frequent fire: regeneration following two large wildfires 12 years apart. Fire Ecology 13(1):91-103. https://doi.org/10.4996/fireecology.1301091
Harrington, C. A., and W. D. Devine. 2006. A practical guide to oak release. General Technical Report PNW-GTR-666. U.S. Forest Service, Pacific Northwest Research Station, Portland, Oregon, USA. https://doi.org/10.2737/PNW-GTR-666
Hastings, M. S., J. S. Barnhart, and R. J. McBride. 1997. Restoration management of northern oak woodlands. Pages 275-280 in Proceedings of a Symposium on oak woodlands: ecology, management, and urban interface issues. General Technical Report PSW-GTR-160. U.S. Forest Service, Pacific Southwest Research Station, Albany, California, USA.
Hayashida, F. M. 2005. Archaeology, ecological history, and conservation. Annual Review of Anthropology 34(1):43-65. https://doi.org/10.1146/annurev.anthro.34.081804.120515
Healy, W. M. 1997. Thinning New England oak stands to enhance acorn production. Northern Journal of Applied Forestry 14(3):152-156. https://doi.org/10.1093/njaf/14.3.152
Healy, W. M., A. M. Lewis, and E. F. Boose. 1999. Variation of red oak acorn production. Forest Ecology and Management 116(1-3):1-11. https://doi.org/10.1016/S0378-1127(98)00460-5
Hilderbrant, W. R. 2007. Northwest California: ancient lifeways. Chapter 7 in T. L. Jones and K. A. Klar, editors. California prehistory: colonization, culture and complexity. Rowman AltaMira, Lanham, Maryland, USA.
Horney, M., R. B. Standiford, D. McCreary, J. Tecklin, and R. Richards. 2002. Effects of wildfire on blue oak in the northern Sacramento Valley. Pages 261-267 in Proceedings of the Fifth Symposium on Oak Woodlands: oaks in California’s challenging landscape. General Technical Report PSW-GTR-184. U.S. Forest Service, Pacific Southwest Research Station, Albany, California, USA.
Huffman, M. R. 2013. The many elements of traditional fire knowledge: synthesis, classification, and aids to cross-cultural problem solving in fire-dependent systems around the world. Ecology and Society 18(4):3. https://doi.org/10.5751/ES-05843-180403
Hunter, J. C., and M. G. Barbour. 2001. Through-growth by Pseudotsuga menziesii: a mechanism for change in forest composition without canopy gap. Journal of Vegetation Science 12:445-452. https://doi.org/10.2307/3236996
Keeley, J. E. 2002. Native American impacts on fire regimes of the California coastal ranges. Journal of Biogeography 29(3):303-320. https://doi.org/10.1046/j.1365-2699.2002.00676.x
Kelly, L. T., and L. Brotons. 2017. Using fire to promote biodiversity. Science 355(6331):1264-1265. https://doi.org/10.1126/science.aam7672
Kimmerer, R. W., and F. K. Lake. 2001. The role of indigenous burning in land management. Journal of Forestry 99(11):36-41.
Kramer, H. A., B. M. Collins, F. K. Lake, M. K. Jakubowski, S. L. Stephens, M. Kelly. 2016. Estimating ladder fuels: a new approach combining field photography with LiDAR. Remote Sensing 8(9):766. https://doi.org/10.3390/rs8090766
Lake, F. K., V. Wright, P. Morgan, M. McFadzen, D. McWethy, and C. Stevens-Rumman. 2017. Returning fire to the land: celebrating traditional knowledge and fire. Journal of Forestry 115(5):343-353. https://doi.org/10.5849/jof.2016-043R2
Lee, G. D. 1998. Walking where we lived: memoirs of a Mono Indian family. University of Oklahoma Press, Norman, Oklahoma, USA.
Lepofsky, D., C. G. Armstrong, S. Greening, J. Jackley, J. Carpenter, B. Guernsey, D. Matthews, and N. J. Turner. 2017. Historical ecology of cultural keystone places of the Northwest Coast. American Anthropologist 119(3):448-463. https://doi.org/10.1111/aman.12893
Lightfoot, K. G., and R. Q. Cuthrell. 2015. Anthropogenic burning and the Anthropocene in late-Holocene California. Holocene 25(10):1581-1587. https://doi.org/10.1177/0959683615588376
Lightfoot, K., R. Cuthrell, C. Boone, R. Byrne, A. Chavez, L. Collins, A. Cowart, R. Evett, P. Fine, D. Gifford-Gonzalez, M. Hylkema, V. Lopez, T. Misiewicz, and R. Reid. 2013. Anthropogenic burning on the central California coast in Late Holocene and early historical times: findings, implications, and future directions. California Archaeology 5:371-390. https://doi.org/10.1179/1947461X13Z.00000000020
Livingston, A. C., J. M. Varner, E. S. Jules, J. M. Kane, and L. A. Arguello. 2016. Prescribed fire and conifer removal promote positive understorey vegetation responses in oak woodlands. Journal of Applied Ecology 53(5):1604-1612. https://doi.org/10.1111/1365-2664.12703
Long, J. W., M. K. Anderson, L. Quinn-Davidson, R. W. Goode, F. K. Lake, and C. N. Skinner. 2016. Restoring California black oak ecosystems to promote tribal values and wildlife. General Technical Report PSW GTR-252. U.S. Forest Service, Pacific Southwest Research Station, Albany, California, USA. https://doi.org/10.2737/PSW-GTR-252
Long, J. W., R. W. Goode, R. J. Gutteriez, J. J. Lackey, and M. K. Anderson. 2017. Managing California black oak for tribal ecocultural restoration. Journal of Forestry 115(5):426-434. https://doi.org/10.5849/jof.16-033
Long, J. W., A. Gray, and F. Lake. 2018. Recent trends in large hardwoods in the Pacific Northwest, USA. Forests 9(10):651. https://doi.org/10.3390/f9100651
Long, J. W., and F. K. Lake. 2018. Escaping social-ecological traps through tribal stewardship on national forest lands in the Pacific Northwest, United States of America. Ecology and Society 23(2):10. https://doi.org/10.5751/ES-10041-230210
Long, J. W., F. K. Lake, R. W. Goode, and M. B. Burnette. 2020. How traditional tribal perspectives influence ecosystem restoration. Ecopsychology 12(2):71-82. https://doi.org/10.1089/eco.2019.0055
Madley, B. 2016. An American genocide: the United States and the California Indian Catastrophe, 1846-1873. Yale University Press, New Haven, Connecticut, USA.
Marcos, G. M., J. J. Obrador, E. Garcia, E. Cubera, M. J. Montero, F. Pulido, and C. Dupraz. 2007. Driving competitive and facilitative interactions in oak dehesas through management practices. Agroforestry Systems 70:25-40. https://doi.org/10.1007/s10457-007-9036-y
McDonald, P. M. 1969. Silvical characteristics of California black oak (Quercus kelloggii Newb.). Forest Service Research Paper PSW-53. U.S. Forest Service PSW-53. Pacific Southwest Forest and Range Experiment Station, Berkeley, California, USA.
McDonald, P. M., R. M. Burns, and B. H. Honkala. 1990. Quercus kelloggii Newb., California black oak. Silvics of North America. Volume 2, Hardwoods. Agriculture Handbook 654:661-671.
McDonald, P. M., and N. R. Vaughn. 2007. Growth of thinned and unthinned hardwood stands on a good site in northern California. General Technical Report PSW-GTR-204. U.S. Forest Service, Pacific Southwest Research Station, Albany, California, USA. https://doi.org/10.2737/PSW-GTR-204
Meentemeyer, R., D. Rizzo, W. Mark, and E. Lotz. 2004. Mapping the risk of establishment and spread of sudden oak death in California. Forest Ecology and Management 200(1-3):195-214. https://doi.org/10.1016/j.foreco.2004.06.021
Miller, A. M., and I. Davidson-Hunt. 2010. Fire, agency and scale in the creation of aboriginal cultural landscapes. Human Ecology 38(3):401-414. https://doi.org/10.1007/s10745-010-9325-3
Milliken, R. 2007. Punctuated culture change in the San Francisco Bay area. Chapter 7 in T. L. Jones and K. A. Klar, editors. California prehistory: colonization, culture and complexity. Rowman AltaMira, Lanham, Maryland, USA.
Minnich, R. A., M. G. Barbour, J. H. Burk, and J. Sosa-Ramírez. 2000. Californian mixed-conifer forests under unmanaged fire regimes in the Sierra San Pedro Mártir, Baja California, Mexico. Journal of Biogeography 27(1):105-129. https://doi.org/10.1046/j.1365-2699.2000.00368.x
Moreno, G., and E. Cubera. 2008. Impact of stand density on water status and leaf gas exchange in Quercus ilex. Forest Ecology and Management 254(1):74-84. https://doi.org/10.1016/j.foreco.2007.07.029
Muller, S. 2003. Towards decolonisation of Australia’s protected area management: the Nantawarrina Indigenous Protected Area experience. Australian Geographical Studies 41(1):29-43. https://doi.org/10.1111/1467-8470.00190
Nemens, D. G., J. M. Varner, and P. W. Dunwiddie. 2019. Resilience of Oregon white oak to reintroduction of fire. Fire Ecology 15:29. https://doi.org/10.1186/s42408-019-0045-9
Nemens, D. G., J. M. Varner, K. R. Kidd, and B. Wing. 2018. Do repeated wildfires promote restoration of oak woodlands in mixed-conifer landscapes? Forest Ecology and Management 427:143-151. https://doi.org/10.1016/j.foreco.2018.05.023
Norgaard, K. M. 2014. The politics of fire and the social impacts of fire exclusion on the Klamath. Humboldt Journal of Social Relations 36:77-101.
Nowacki, G. J., and M. D. Abrams. 2008. The demise of fire and “mesophication” of forests in the eastern United States. BioScience 58(2):123-138. https://doi.org/10.1641/B580207
Oheimb, G. V., A. C. Lang, H. Bruelheide, D. I. Forrester, I. Wäsche, M. Yu, and W. Härdtle. 2011. Individual-tree radial growth in a subtropical broad-leaved forest: the role of local neighbourhood competition. Forest Ecology Management 261(3):499-507. https://doi.org/10.1016/j.foreco.2010.10.035
Pearce, T., J. Ford, A. C. Willox, and B. Smit. 2015. Inuit traditional ecological knowledge (TEK), subsistence hunting and adaptation to climate change in the Canadian Arctic. Arctic 68(2):233-245. https://doi.org/10.14430/arctic4475
Quinn-Davidson, L. N., and J. M. Varner. 2012. Impediments to prescribed fire across agency, landscape, and manager: an example from northern California. International Journal of Wildland Fire 21(3):210-218. https://doi.org/10.1071/WF11017
Robinson, C. J., K. Maclean, R. Hill, E. Bock, and P. Rist. 2016. Participatory mapping to negotiate indigenous knowledge used to assess environmental risk. Sustainability Science 11(1):115-126. https://doi.org/10.1007/s11625-015-0292-x
Rose, A. K., C. H. Greenberg, and T. M. Fearer. 2012. Acorn production prediction models for five common oak species of the eastern United States. Journal of Wildlife Management 76(4):750-758. https://doi.org/10.1002/jwmg.291
Schriver, M., R. L. Sherriff, J. M. Varner, L. Quinn-Davidson, and Y. Valachovic. 2018. Age and stand structure of oak woodlands along a gradient of conifer encroachment in northwestern California. Ecosphere 9(10):e02446. https://doi.org/10.1002/ecs2.2446
Shebitz, D. J., S. H. Reichard, and P. W. Dunwiddie. 2009. Ecological and cultural significance of burning beargrass habitat on the Olympic Peninsula, Washington. Ecological Restoration 27(3):306-319. https://doi.org/10.3368/er.27.3.306
Skinner, C. N., A. H. Taylor, and J. K. Agee. 2006. Klamath Mountains Bioregion. Pages 170-194 in N. G. Sugihara, J. W. van Wagtendonk, J. Fites-Kaufman, K. E. Shaffer, and A. E. Thode, editors. Fire in California’s ecosystems. University of California Press, Berkeley, California, USA. https://doi.org/10.1525/california/9780520246058.003.0009
Spector, T., and F. E. Putz. 2006. Crown retreat of open-grown Southern live oaks (Quercus virginiana) due to canopy encroachment in Florida, USA. Forest Ecology and Management 228(1-3):168-176. https://doi.org/10.1016/j.foreco.2006.03.001
Stephens, S. L., R. E. Martin, and N. E. Clinton. 2007. Prehistoric fire area and emissions from California’s forests, woodlands, shrublands, and grasslands. Forest Ecology and Management 251(3):205-216. https://doi.org/10.1016/j.foreco.2007.06.005
Stephens, S. L., J. J. Moghaddas, C. Edminster, C. E. Fiedler, S. Haase, M. Harrington, J. E. Keeley, E. E. Knapp, J. D. McIver, K. Metlen, C. N. Skinner, and A. Youngblood. 2009. Fire treatment effects on vegetation structure, fuels, and potential fire severity in western U.S. forests. Ecological Applications 19(2):305-320. https://doi.org/10.1890/07-1755.1
Wainwright, T. C., and M. G. Barbour. 1984. Characteristics of mixed evergreen forest in the Sonoma Mountains of California. Madroño 31(4):219-230.
Werth, P. A., B. E. Potter, C. B. Clements, M. A. Finney, S. L. Goodrick, M. E. Alexander, M. G. Cruz, J. A. Forthofer, and S. S. McAllister. 2011. Synthesis of knowledge of extreme fire behavior: volume I for fire managers. General Technical Report PNW-GTR-854. U.S. Forest Service, Pacific Northwest Research Station, Portland, Oregon, USA. https://doi.org/10.2737/PNW-GTR-854
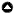
Fig. 1

Fig. 1. Black oak trees (Quercus kelloggii) at Pepperwood Preserve after a wildfire. From left to right; on the edge of grassland, with encroaching Douglas-firs (Pseudotsuga menziesii), landscape view.
Fig. 2

Fig. 2. Maps of the specimen oaks identified at Pepperwood Preserve with respect to (a) local species alliances and (b) fire severity during the 2019 Kincade Fire. Sentinel-2 data was used to extract the relativized burn ratio (RBR) from the Kincade fires to represent burn severity.
Fig. 3

Fig. 3. (A) The effect of mean diameter at breast height (DBH) of surrounding trees on specimen oak crown area (F1, 51 = 7.33, p = 0.009); (B) The effect of the number of surrounding trees on specimen oak crown area (F1, 51 = 7.58, p = 0.008); and (C) The effect of the interaction between mean DBH of surrounding trees and the number of surrounding trees on specimen oak crown area (F1, 51 = 6.27, p = 0.016).