






The following is the established format for referencing this article:
Londe, D. W., S. M. Cady, R. D. Elmore, and S. D. Fuhlendorf. 2022. Woody plant encroachment pervasive across three socially and ecologically diverse ecoregions. Ecology and Society 27(3):11.ABSTRACT
Woody plant encroachment in grasslands represents one of the greatest challenges for global biodiversity conservation. Furthermore, this is a social-ecological problem, where human activity and behavior have resulted in significant changes in ecological processes that control woody plants, and failure to fully recognize the role of human activity has led to continued loss of grasslands worldwide. It is therefore critical that conservation professionals understand how ecological systems, settlement patterns, and fragmentation from anthropogenic development interact to influence rates of woody plant encroachment. Using annual estimates of tree cover derived from regionally available remote-sensing data, our objectives were, first, to describe rates of woody plant encroachment over the last 20 years (2000–2019) across three ecologically and socially diverse ecoregions in the Southern Great Plains of North America. Then, we examined how anthropogenic and biophysical variables influenced rates of encroachment (both directly and indirectly) within the region. Results indicate that, despite marked differences in social and ecological characteristics, all three ecoregions have experienced consistent increases in woody plant encroachment during the study period. This included the Flint Hills ecoregion of Kansas and Oklahoma, an area that experiences widespread and frequent fires. At the regional scale, rates of encroachment were directly and negatively related to the average area burned in a county, initial cover of trees, and fragmentation from row crops. Percent cover of development and row crop agriculture also indirectly alter rates of encroachment in a county by influencing initial tree cover and fire activity. The pervasive nature of woody plant encroachment, even in regions that experience frequent fires, suggests that many grasslands are being managed outside of critical ecological thresholds needed to maintain grasslands and limit woody encroachment, which can have significant implications for biodiversity. Our results show that anthropogenic or ecological factors do not act in isolation in their influence on woody plant encroachment and can form complex relationships that shape regional trends in woody plant encroachment.INTRODUCTION
Grassland and savannah ecosystems are undergoing a rapid change from plant communities dominated by herbaceous species to a new state dominated by woody plants (Roques et al. 2001, Briggs et al. 2005, Stevens et al. 2017). This regime shift, termed woody plant encroachment, is a global challenge affecting grasslands and savannahs across multiple continents and has significant implications for global biodiversity (Stevens et al. 2017, Venter et al. 2018, García Criado et al. 2020). Woody plant encroachment has been partly attributed to widespread suppression of fire by humans, which has allowed disturbance-sensitive tree species to increase in coverage and extent in remaining grasslands (Sholes and Archer 1997, Van Auken 2000, Bond and Keeley 2005, Briggs et al. 2005, Archer et al. 2017). Factors such as rising CO₂ levels (Bond and Midgley 2000), increases in nitrogen (Kochy and Wilson 2001), and overgrazing of grasslands have also been linked to accelerating tree encroachment (Archer et al. 2017). These changes in grassland systems can lead to a loss in biodiversity (Ratajczak et al. 2012) and in productivity of forage (Van Auken 2009), reduced ecological resilience (Alofs and Fowler 2013, Eldridge et al. 2015), interrupted ecosystem services (Archer et al. 2001, Eldridge et al. 2015, Koch et al. 2015), and loss of habitat for grassland dependent species (Grant et al. 2004, Fuhlendorf et al. 2017, Lautenbach et al. 2017). Because grasslands and savannahs worldwide have already undergone extensive reduction and degradation from anthropogenic activity (Hoekstra et al. 2005), the increase in woody plant cover poses an existential conservation challenge for remaining grasslands. Understanding the causes and scope of woody plant encroachment is a vital step toward effectively addressing this challenge.
An emerging paradigm in grassland conservation is the shift toward understanding that woody plant encroachment in grasslands is a social-ecological problem (Donovan et al. 2018, Luvuno et al. 2018, Wilcox et al. 2018). Under this paradigm, it is understood that human activity and behavior interacts with underlying ecological patterns to determine the maintenance of a landscape in either a grassland or woodland state. At broad scales, woody plant encroachment is controlled by both the social and economic factors that determine whether a region implements effective control measures for woody plants, such as frequent fires or targeted mechanical/herbicide treatments (Wilcox et al. 2018). These broad patterns are influenced by historical land-use practices (Higgins et al. 1999), local regulations regarding land use and liability associated with use of prescribed fire (Yoder et al. 2004, Kreuter et al. 2019), capital or resource constraints, and cultural or personal perceptions about the danger or utility of fire (Harr et al. 2014, Hoffman et al. 2021). Other factors relating to human activity such as settlement patterns, alternative land uses, and intensity of fragmentation can further influence tree encroachment by creating refuges where trees can become established or by altering disturbance patterns, such as fire and grazing, that would have otherwise limited encroachment (Hoch and Briggs 1999, Briggs et al. 2005, Scholtz et al. 2018a), or through the intentional planting of trees (Gardner 2009). However, ecological factors such as rainfall (Scholtz et al. 2018b), soils (Kaur et al. 2020), and distance to seed sources (Donovan et al. 2018), place limits on where trees could become established. Social and ecological factors can further interact with each other to form feedback loops that further reinforce patterns of encroachment (Ratajczak et al. 2012, Luvuno et al. 2018). Despite these known or perceived interactions between social and ecological landscape patterns in shaping the distribution of woody plants, most studies treat these factors as independent processes and rarely consider them as part of a causal network that can form complex interactions and feedbacks.
The Southern Great Plains of North America has experienced encroachment at rates five to seven times higher than any other region in North America, threatening much of the remaining grasslands in the region (Engle et al. 2008, Barger et al. 2011). However, the Southern Great Plains is a culturally and ecologically diverse region, which has resulted in the many imbedded ecoregions having experienced different rates of encroachment over the last century (Barger et al. 2011). An east-west precipitation gradient and a diversity of soil types largely determine productivity of grasslands in the region, and in turn settlement patterns and land uses by European settlers. The resulting differences in cultural and economic practices set many ecoregions on different trajectories in terms of woody plant encroachment and responses (Wilcox et al. 2018). For example, widespread fire suppression, improper grazing, and fragmentation from row crop agriculture has facilitated encroachment in the southwestern extent of the region in Texas (Box 1967, Diamond and True 2008). In contrast, shallow rocky soils that prevented cultivation and a widespread and persistent fire culture in the Flint Hills ecoregion have allowed grasslands to persist with low levels of encroachment (Hoy 1989, Briggs et al. 2005), though recent studies suggest woody plant encroachment may be an emerging problem in the region (Hoch and Briggs 1999, Briggs et al. 2005, Ratajzak et al. 2016). Because of the diverse nature of the region, the Southern Great Plains represents a potentially important case study for understanding how social and ecological factors interact to influence rates of woody plant encroachment.
Increasing emphasis is being placed on understanding woody plant encroachment as a social-ecological challenge. Developing a clear model for the complex interactions between anthropogenic land-use change, management, and rates of woody plant encroachment is needed to better understand the drivers of this global conversion. Our objective in this study was to describe trends in woody plant encroachment across three ecoregions in the Southern Great Plains of North America that differ in their social and ecological composition. Specifically, we tested the hypothesis that ecoregions where broad socioeconomic patterns favor widespread and frequent fire as a management tool would experience limited woody plant encroachment, whereas ecoregions where the prevailing cultural attitudes toward fire preclude effective management practices would have the highest rates of encroachment. Our secondary objective was to assess the causal relationships between regional anthropogenic and ecological factors and woody plant encroachment and determine how these factors interact to shape patterns of woody plant encroachment. To match the scale of our first objective, we used variables related to land use, development, and fragmentation as proxies for differences in regional social and anthropogenic activities, and variables related to fire and availability of seed sources to represent ecological processes. Although many studies exist documenting increases in woody plant cover throughout the Great Plains and local scale anthropogenic and ecological factors leading to this rise in woody plants, few studies exist that examine the factors that influence woody plant encroachment at broad regional scales. By examining how landscape level variables influence the rate of woody plant encroachment across several ecoregions that make up gradients of ecological, anthropogenic, and biophysical variables, we attempt to provide a framework for understanding woody plant encroachment in the Great Plains, with this knowledge being able to guide conservation decisions at the regional scale.
STUDY SITE
To investigate rates of woody plant encroachment and the factors that influence encroachment we selected three ecoregions described in Wilcox et al. (2018) as a case study. These ecoregions, the Edwards Plateau of Texas, the Flint Hills of Kansas and Oklahoma, and the Rolling Red Hills of western Oklahoma were selected to represent gradients in ecological and anthropogenic variables that are predicted to influence rates of woody plant encroachment (Table 1, Fig. 1; Wilcox et al. 2018). The three ecoregions fall along an east-west rainfall gradient, ranging from a mean annual precipitation of around 900 mm in the eastern Flint Hills to 670 mm in the Rolling Red Hills. This rainfall gradient results in a corresponding gradient of productivity ranging from tallgrass prairie communities in the east to short-grass vegetation communities in the west (Bragg and Streuter 1996, Assal et al. 2015). All three regions were dominated by grassland vegetation communities with scattered woodlands and forests that persisted in areas with limited fire activity (Wells, 1965, Fowler and Dunlap 1986, Bragg and Streuter 1996). Following European settlement, land use within each region was dominated by cattle grazing until the early 1900s, when increased settlement resulted in a shift toward crop production in many areas (Wilcox et al. 2018). The Flint Hills is the exception to this trend of increased crop production, because shallow rocky soils prohibited widespread tillage in the region. Use of fire has largely persisted in the Flint Hills primarily as a management tool for removing old grass litter and stimulating new grass growth (Hoy 1989, Briggs et al. 2005). Use of fire as a management tool is much rarer in the Rolling Red Hills and Edwards Plateau (Table 1). Though much of the three ecoregions is still primarily used for pastoral activities and agricultural activities, these regions have also undergone considerable development for human settlements (Table 1). All three ecoregions have similar amounts of total cover for development but show diverging trends in population density and road density (Table 1). The Edwards Plateau contains several metropolitan areas (including San Antonio and Austin, TX) resulting in a higher average population density, although the Rolling Red Hills has remained relatively sparsely populated. The Flint Hills is intermediate in terms of population compared to the other two regions, with several large cities (e.g., Manhattan, KS) interspersed with open rangelands (Hoch and Briggs 1999). Compared to the other two regions, the Flint Hills has a higher overall density of roads per county.
METHODS
Data preparation
Rates of encroachment
To define the extent of the three ecoregions considered, we used Level III U.S. Environmental Protection Agency ecoregions to define the Edwards Plateau and the Flint Hills, and Level IV ecoregions to define the Rolling Red Hills. To delineate a smaller, regional scale, we divided each of the three ecoregions into counties (Edwards Plateau encompassed 36 counties, Flint Hills encompassed 20 counties, and Rolling Red Hills encompassed nine counties). To estimate woody plant encroachment, we used data available from the Rangeland Analysis Platform, an annual resolution, 30 m grain, plant functional type dataset (Jones et al. 2018; https://rangelands.app/]). For each county, we downloaded the time-series data for tree cover from 2000 to 2019. To estimate the average annual rate of change in tree cover for each county, we used simple linear regression where the year (2000–2019) was used as the predictor variable and the estimated tree cover for each year was the response variable. The estimated slope was interpreted as the average annual rate of change per year for each county where a positive slope represents increasing woody plant encroachment, and a negative slope represents woody plant decline. Although linear regression does not allow for the detection of nonlinear relationships and short-term fluctuations in tree cover, the use of relatively simple models allowed us to estimate broad trends in tree cover for the study period while minimizing the influence of variance inherent in remotely sensed data.
Structural equation model development and anthropogenic-ecological variables
We developed predictions that would serve as the foundation for the structural equation models through a review of recent literature that assessed different factors associated with woody plant or invasive species encroachment (Table 2). Because of the course resolution of the woody plant data, we focused on developing hypotheses that could be represented by broad-scale landscape variables such as cover of different land uses, population density, and road density so that the resolution of our social-ecological data matched the resolution of the woody plant data.
For the anthropogenic and ecological variables, we downloaded county-level data from freely available online data sources for each region we investigated (Table 1). Because our objective was to determine the relative importance of anthropogenic versus ecological patterns on woody plant encroachment, we chose to focus on variables from categories that have been shown to influence woody plant encroachment in past studies (Table 1). We calculated population density for each county as the number of people counted in the 2000 Census divided by the area of county (km²). We calculated road density using the national TIGER road database and calculated the length of road per km² in each county. We used data from the Rangeland Analysis Platform to estimate tree and grassland cover in each county at the start of the study period. Because the Rangeland Analysis platform does not provide estimates of non-vegetation landcover we used the 2011 National Land Cover data set to estimate percent cover of agriculture and development (Scholtz et al. 2018a). To find the mean annual precipitation for each county we used 4-km gridded national weather maps from the Climate PRISM group, using the long-term average datasets (https://prism.oregonstate.edu/normals/). For fire data, we downloaded fire perimeter shapefiles from the Monitoring Trends in Burn Severity (MTBS) database (Eidenshink et al. 2007, Picotte et al. 2020). MTBS data have a minimum mapping resolution of 404 ha, so our results may underestimate fire activity because many small fires may not be represented. However, because we are attempting to develop an index of fire activity during our study period rather than map individual fires, we believe MTBS data represent the best option for estimating fire activity over large geographic areas. We combined all fires in each county into a single fire variable because 75% of the fires recorded in our study area were classified as having unknown origins. This precluded us from developing any hypothesis about the role of prescribed fire versus wildfire in our analysis. We then estimated the average proportion of the county burned each year for the period of 2000 to 2019. We converted burned areas to proportion of counties burned to standardize our measure of fire activity across counties of different sizes. To verify that our choice of datasets did not influence the overall findings of our study we calculated the same variable using the MODIS burned area product (Roy et al. 2005). We found the Pearson’s correlation between the average area estimated using the MTBS dataset and the MODIS dataset was rs = 0.99. This suggests there is considerable overlap in datasets, and that both datasets are largely detecting the same fires over the course of our study. All data management and processing were conducted in ArcGIS Pro (ESRI, Redlands, CA).
Data Analysis
Ecoregion comparisons
To compare rates of encroachment in the three ecoregions we used the Kruskal-Wallis test. We then used a Dunn test to perform a post-hoc test to determine pairwise differences among the ecoregions. All tests were considered significant at the p < 0.05 level.
Anthropogenic-biophysical variables
We assessed the influence of social-ecological factors on woody plant encroachment using a structural equation modeling approach. As the foundation for our analysis, we developed a path diagram representing connections predicted by the hypothesis presented in Table 2. We then assessed the strength of relationships between variables using maximum likelihood methods using the R Package “lavaan” (Rosseel 2012). We did not attempt to reduce the original model, because we were testing a specific hypothesis about the relationship of different landscape variables and rates of woody plant encroachment. We did calculate modification indices to determine if there were any missing causal links. This resulted in the addition of a single link between mean annual precipitation and initial cover of trees in a county. We evaluated the final fit of the model using a χ² test and reporting the degrees of freedom and root mean square error (RMSE).
RESULTS
Analysis of land-cover data showed a trend of increasing tree cover in all three ecoregions (Fig. 2; Figs. A1, A2, A3, Appendix 1). The Edwards Plateau showed the lowest overall percent change in tree cover with a 18.62% increase from 2000 to 2019 (Table 3). The Flint Hills ecoregion had the highest percent change in tree cover at a 46.6%, and the Rolling Red Hills ecoregion was intermediate at a 39.2%. As a result of differing amounts of initial tree cover and the size of the ecoregions, this resulted in the conversion of 372,930 ha, 39,896 ha, and 84,232 ha from grasslands to woody plants in the Edwards Plateau, Rolling Red Hills, and Flint Hills ecoregions, respectively (Table 3). Despite the differences in overall area converted to trees, the mean rate of encroachment at the county level did not differ significantly between the three ecoregions (χ² = 5.00, p = 0.08), but this may have been in part because of the considerable within ecoregion variability in woody plant encroachment (Fig. 2; Figs. A1, A2, A3, Appendix 1).Structural equation models
Fit indices indicated mixed evidence for the overall fit of our model. The Standardized Root Mean Square Residual (SRMSR) and Comparative Fit Index (CFI) indicated a good fit (SRMSR = 0.048, CFI = 0.96; lower values for SRMSR and values close to one for CFI indicate a good fit) whereas high values of Root Mean Square Error (RMSE) indicated a large amount of variance remained unexplained (RMSE = 0.187; values less than 0.06 are considered good; Fan et al. 2016). Our model indicated that many of our variables influenced woody plant encroachment both directly and indirectly (Fig. 3). Counties with greater areas that were burned annually had lower overall rates of woody plant encroachment supporting Hypothesis 1, that greater fire activity would limit tree encroachment (β = -0.07, SE = 0.02, p = 0.004). In contrast to Hypothesis 2, which predicted counties with greater tree cover would have higher rates of encroachment, initial cover of trees in each county had a negative effect on subsequent rates of encroachment (β = -0.08, SE = 0.03, p = 0.01). However, the observed relationship between initial tree cover and rate of encroachment, and the negative relationship between encroachment and percent crop in a county (β = -0.07, SE = 0.03, p = 0.01), provide support for Hypothesis 3, that counties with greater amounts of non-grassland landcover would experience lower rates of encroachment. Finally, counties with higher mean annual precipitation had higher rates of encroachment supporting Hypothesis 4 (β = 0.10, SE = 0.04, p = 0.006). Road density and the percent cover of development in a county had nonsignificant direct effects on woody plant encroachment (Fig. 3).
We also observed several indirect relationships between anthropogenic variables and woody plant encroachment. Specifically, counties with greater fragmentation from alternative land cover types such as roads (β = -0.23, SE = 0.15, p = 0.1) or development (β = -1.11, SE = 0.46, p = 0.02) in general had lower overall areas burned annually, whereas road density had a positive relationship with area burned (β = 1.24, SE = 0.27, p < 0.001). This provides mixed support for Hypothesis 5, which states that counties with greater fragmentation would have less fire activity. We found mixed support for the hypothesis that increased fragmentation would influence initial tree cover (Hypothesis 6), because percent agriculture (β = -0.60, SE = 0.09, p < 0.001) and road density (β = -0.79, SE = 0.29, p = 0.10) had a negative effect on initial tree cover whereas percent developed (β = 0.25, SE = 0.32, p = 0.36) had a nonsignificant effect on initial tree cover. Population density in a county did not influence initial tree cover (β = 0.34, SE = 0.24, p = 0.15) or average area burned (β = -0.04, SE = 0.41, p = 0.92) in a county, suggesting no support for Hypothesis 7.
DISCUSSION
Historically, factors such as precipitation, soils, and disturbance from herbivory and fires (many of which were anthropogenic, particularly in the Great Plains) limited the ability of woody plants to spread and become established in grasslands (Archer et al. 2017). Because human populations have expanded and are becoming an increasingly dominant force in shaping biotic communities worldwide (Ellis and Ramankutty 2008), changes in human activity have played an important role in facilitating the rise in woody plants in grasslands across the globe (Archer et al. 2017, Stevens et al. 2017, Venter et al. 2018). Here, we found that three grassland-dominated ecoregions, which differ in their social and ecological characteristics, are currently experiencing widespread woody plant encroachment. Most notable, the Flint Hills ecoregion, where prescribed fire is a common grassland management tool, had rates of encroachment that were comparable to ecoregions that receive limited fire. Further, our results show that anthropogenic and ecological factors rarely act in isolation in their influence on woody plant encroachment and can form complex relationships that shape regional trends in woody plant encroachment. Grasslands are among the most heavily altered ecosystems in the world (Hoekstra et al. 2005), and the pervasive nature of woody plant encroachment represents a significant conservation challenge that current management practices and strategies may not be sufficient to address (Fogarty et al. 2020, Scholtz et al. 2021). Further, continued encroachment has the potential to alter a number of important ecosystem services provided by remaining grasslands (Archer and Predick 2014). Failure to account for the direct and indirect relationships between landscape variables and woody plant encroachment can hinder our ability to accurately predict trends in encroachment.
By generating multiple hypotheses and developing a causal diagram describing the direct and indirect effects of anthropogenic and ecological factors, we were better able to elucidate the role these variables had in shaping woody plant encroachment across the three ecoregions. Our predictions that woody plant encroachment would be higher in counties with greater annual precipitation (Sankaran et al. 2005, Scholtz et al. 2018b), and lower in areas with greater fire activity (Archer et al. 2017) were generally corroborated. However, we found that several of the remaining variables that were considered had more complex relationships with woody plant encroachment. Some of the anthropogenic variables, such as roads and development, appear to influence woody plant encroachment indirectly through other variables such as area burned, whereas other anthropogenic factors, such as fragmentation from crops, influenced woody plants both directly and indirectly. Although this supports previous research indicating woody plant encroachment is higher in areas with greater fragmentation (Scholtz 2018a), these results suggest consideration of the specific source or type of fragmentation is important because different landscape factors can interact with existing ecological processes in a variety of ways. In contrast to our stated hypothesis that increased tree cover in a county would result in greater propagule pressure and thus higher rates of encroachment, we found rates of encroachment were lower in counties with high tree cover. Lack of support for increased tree cover leading to higher rates of invasion in the surrounding grasslands may be in part because we could not distinguish between different tree species found in the region: different species can vary considerably in seed production rates and invasive ability (Richardson et al. 1994). Although consideration of propagule source and pressure may be important for assessing local scale probability of encroachment (Lockwood et al 2005), our results suggest this relationship may be more complex at regional scales. Factors such as rainfall, woody plant species, and available space for new trees to invade may also be important considerations.
Despite the growing acknowledgement of woody plant encroachment as a social-ecological process (Donovan et al. 2018, Luvuno et al. 2018, Wilcox et al. 2018), these factors are often studied separately. Human behavior and anthropogenic modifications to the landscape can affect woody plants directly either through afforestation practices or tree removal (Gardner 2009, Ding and Eldridge 2019), or indirectly by alterations in the ecological processes that would have otherwise influenced tree encroachment. Changes in the extent and frequency of fire has been cited as an important driver in the rise of woody plants globally (Bond and Keely 2005, Archer et al. 2017). Although there are many factors that can influence the use of fire as a management tool, our study indicates that increased fragmentation from development and row crops had a negative relationship with average area burned in a county during the study period, which then potentially facilitated woody plant encroachment. This reduction in fire may be the result of changes in the ability of fire to spread across the landscape, or because of reduced fires, or use of low intensity fires when burning near developed areas (Pickens et al. 2017). In contrast, rates of encroachment were also lower in counties with greater fragmentation, particularly from agriculture, potentially because of reduced grasslands for trees to invade and become established. These contrasting relationships between fragmentation and woody plant encroachment highlight the importance of studying social and ecological factors together, because considering the effects of fragmentation alone would have likely resulted in an incomplete understanding of the effects of landscape change on woody plant encroachment.
Despite differences in management practices and land use, all three ecoregions had similar rates of woody encroachment. However, these similarities may be superficial, and more accurately represent different stages of the invasion process. Many biological invasions follow a consistent pattern, where initial establishment and population growth may be slow because of limited propagule sources or interactions with already present species, but then exhibit periods of rapid population growth once a critical population size is reached (Crooks 2005), followed finally by a slowing of growth as a population approaches the carrying capacity of a landscape (Shigesada and Kawasaki 1997). Whereas all three ecoregions were historically dominated by grassland vegetation, different histories of settlement and land use in these ecoregions have caused these regions to have undergone different levels of tree invasion prior to the time period of our study. The persistent use of prescribed fire as a management tool in the Flint Hills has likely maintained this region in the early stages of invasion, where spread of trees is slow as new individuals struggle to become established as a result of frequent burning (Hoy 1989, Briggs et al. 2005). Alternatively, the Edwards Plateau may represent the endpoint of a biological invasion, where encroachment may be slowing because of the higher number of established trees and higher levels of grassland fragmentation in the region limits the available space for new trees to become established (Archer et al. 2017). Importantly, despite current similarities in rates of encroachment, subsequent population dynamics of invading trees may be very different for regions at different stages of invasion, and management strategies and resources should be allocated in anticipation of these potential changes in population growth (Yokomizo et al. 2009, Roberts et al. 2018). For example, management in the Flint Hills should place greater emphasis on preventative measures and targeted removal of trees in key areas needed for maintaining grassland connectivity in the region, whereas in the Edwards Plateau more intensive management may be needed to remove trees and management should be targeted to areas with the greatest restoration potential (Yokomizo et al. 2009, Roberts et al. 2018).
Despite the negative relationship between encroachment and the average area burned annually in a county, the Flint Hills ecoregion, where a widespread fire culture has largely persisted, was shown to be experiencing rates of encroachment similar to regions with little to no fire. Recent studies of the spatial and temporal patterns of fires in the Flint Hills showed considerable variation among landowners in terms of the frequency with which fire was applied to the landscape (Ratajzak et al. 2016). Although many landowners burn frequently, as much as 57% of grasslands in the Flint Hills experiences a fire frequency that is too low to prevent woody plant encroachment, with at least one fire every one to three years (Ratajzak et al. 2016). Individual landowners or agencies may exhibit a wide array of behaviors as a result of differing land uses for their property (Coppedge et al. 2001), personal ethics, risk perception for different management tools (Toledo et al. 2013, Harr et al. 2014), aesthetic preferences (Sharp et al. 2012, Stroman et al. 2020), as well as capital or resource constraints that influence how they manage their property (Toledo et al. 2014). Additionally, many managers have changed when they burn during the year to periods when conditions are milder (typically March to April) resulting in lower intensity fires (Twidwell et al. 2016a). These milder burning conditions may result in fires that lack the intensity needed to effectively control woody plants (Twidwell et al. 2016b, Twidwell et al. 2019). Alternatively, the pervasive nature of woody plant encroachment, even in regions where fire is still used as a management tool, has been cited as evidence for global drivers of woody plant encroachment (Bond and Midgley 2000, Stevens et al. 2017, García Criado et al. 2020). For example, increasing atmospheric CO₂ and warmer temperatures that are the result of climate change, may provide a competitive advantage to woody plants that use C₃ photosynthetic pathways over herbaceous C₄ species that dominate in many grasslands (Bond and Midgley 2000). A critical step forward in understanding the causes and consequences of human activities on woody plant encroachment will involve investigating the relative importance of top-down (e.g., global climate change) and bottom-up (e.g., local effects of fire or fragmentation) drivers and how these drivers interact to influence encroachment.
Our results indicate that as a whole, managers are losing the battle with woody plants. This is despite decades of brush management, millions of dollars in cost-share programs, and the collective efforts of thousands of managers (Scholtz et al. 2021). This problem transcends regions and the scale of this challenge suggests managing woody plants may require a reevaluation of how and where management takes place. Given the short-lived nature of many efforts to control woody plants, especially in areas where these species may already be common (Fogarty et al. 2021, Scholtz et al. 2021), resources may be better allocated to regions or localities that have low levels of tree cover but are at higher risk of encroachment or have especially high conservation importance to prevent further losses (Yokomizo et al. 2009, Sullins et al. 2019). In order to achieve this, managers must understand the landscape-scale processes that influence woody plant encroachment, and develop a framework that incorporates both anthropogenic and ecological processes for identifying at-risk areas. Our work attempts to begin to address the need. Although local processes are certainly critical for determining the exact rate and extent of woody plant spread, broad-scale analyses such as ours are essential for guiding regional planning.
The loss of grasslands at a global scale has been described as an impending “biome crisis” (Hoekstra et al. 2005) and developing models that acknowledge the diverse and dynamic nature of North American grasslands are essential for effectively allocating conservation and management resources (Wilcox et al. 2018). Confronting this will require a reevaluation of how we understand the social and ecological factors that influence woody plant encroachment, and how we apply management practices to the landscape (Twidwell et al. 2013). Current low amounts of tree cover in ecoregions such as the Flint Hills and the Rolling Red Hills may still allow for early intervention practices, such as increased use of fire, herbicide, or targeted use of mechanical removal. Despite our broad-scale findings, variation in regional trends in encroachment suggest local landscape factors still have an important role in determining encroachment (Higgins and Scheiter 2012, Stevens et al. 2017, Venter et al. 2018) and individual properties can maintain grasslands and grassland biota. Although we distinguished between anthropogenic and ecological variables in this study, in many cases variables such as fire and tree cover cannot be so clearly defined as separate from human behavior, and settlement patterns play a critical role in determining how and where trees and fire occur (Berg et al. 2015). Consideration of woody plant encroachment and human behavior together will be critical for effective management (Wilcox et al. 2018). However, positive feedback loops that are associated with plant community shifts may soon accelerate the process of woody plant encroachment, challenging our ability to manage with current tools and potentially resulting in irreversible losses in biodiversity.
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.AUTHOR CONTRIBUTIONS
D. W. L., S. M. C., R. D. E., and S. D. F. designed and conceived the study. D. W. L. and S. M. C. performed data analysis.
D. W. L. and S. M. C. wrote the manuscript. D. W. L., S. M. C., R. D. E., and S. D. F. provided feedback and comments on all stages of manuscript preparation.
ACKNOWLEDGMENTS
This work is supported by Hatch OKLO3056 project from the United States Department of Agriculture’s National Institute of Food and Agriculture and the Oklahoma Agricultural Experiment Station at Oklahoma State University. We received additional support from the Bollenbach and Groendyke Endowed Wildlife Chairs.
DATA AVAILABILITY
All data used are freely available via online databases described in this paper.
LITERATURE CITED
Alofs, K. M., and N. L. Fowler. 2013. Loss of native herbaceous species due to woody plant encroachment facilitates the establishment of an invasive grass. Ecology 94(3):751-760. https://doi.org/10.1890/12-0732.1
Archer, S. R., E. M. Andersen, K. I. Predick, S. Schwinning, R. J. Steidl, and S. R. Woods. 2017. Woody plant encroachment: causes and consequences. Pages 25-84 in David D. Briske, editor. Rangeland systems: processes, management, and challenges. Springer International, Cham, Switzerland. https://doi.org/10.1007/978-3-319-46709-2_2
Archer, S. R., T. W. Boutton, and K. A. Hibbard. 2001. Trees in grasslands: biogeochemical consequences of woody plant expansion. Pages 115-137 in E.-D. Schulze, S. P. Harrison, M. Heimann, E. A. Holland, J. Lloyd, I. C. Prentice, and D. Schimel, editors. Global biogeochemical cycles in the climate system. Academic, San Diego, California, USA.
Archer, S. R., and K. I. Predick. 2014. An ecosystem services perspective on brush management: research priorities for competing land‐use objectives. Journal of Ecology 102(6):1394-1407. https://doi.org/10.1111/1365-2745.12314
Assal, T. J., C. P. Melcher, and N. B. Carr. 2015. Southern Great Plains rapid ecoregional assessment: pre-assessment report. Report 2331-1258, U.S. Geological Survey, Reston, Virginia, USA. https://doi.org/10.3133/ofr20151003
Barger, N. N., S. R. Archer, J. L. Campbell, C. Y. Huang, J. A. Morton, and A. K. Knapp. 2011. Woody plant proliferation in North American drylands: a synthesis of impacts on ecosystem carbon balance. Geophysical Research: Biogeosciences 116(G4). https://doi.org/10.1029/2010JG001506
Berg, M. D., M. G. Sorice, B. P. Wilcox, J. P. Angerer, E. C. Rhodes, and W. E. Fox. 2015. Demographic changes drive woody plant cover trends—an example from the Great Plains. Rangeland Ecology & Management 68(4):315-321. https://doi.org/10.1016/j.rama.2015.05.004
Bond, W. J., and J. E. Keeley. 2005. Fire as a global ‘herbivore’: the ecology and evolution of flammable ecosystems. Trends in Ecology & Evolution 20(7):387-394. https://doi.org/10.1016/j.tree.2005.04.025
Bond, W. J., and G. F. Midgley. 2000. A proposed CO₂-controlled mechanism of woody plant invasion in grasslands and savannas. Global Change Biology 6(8):865-869. https://doi.org/10.1046/j.1365-2486.2000.00365.x
Box, T. W. 1967. Range deterioration in west Texas. Southwestern Historical Quarterly 71:37-45.
Bragg, T. B., and A. A. Steuter. 1996. Prairie ecology—the mixed prairie. Pages 53-66 in F. B. Samson and F. L. Knopf, editors. Prairie conservation: preserving North America’s most endangered ecosystem. Island, Washington, D.C., USA.
Briggs, J. M., A. K. Knapp, J. M. Blair, J. L. Heisler, G. A. Hoch, M. S. Lett, and J. K. McCarron. 2005. An ecosystem in transition: causes and consequences of the conversion of mesic grassland to shrubland. BioScience 55(3):243-254. https://doi.org/10.1641/0006-3568(2005)055[0243:AEITCA]2.0.CO;2
Coppedge, B. R., D. M. Engle, S. D. Fuhlendorf, R. E. Masters, and M. S. Gregory. 2001. Landscape cover type and pattern dynamics in fragmented Southern Great Plains grasslands, USA. Landscape Ecology 16:677-690. https://doi.org/10.1023/A:1014495526696
Crooks, J. A. 2005. Lag times and exotic species: the ecology and management of biological invasions in slow-motion. Ecoscience 12:316-329. https://doi.org/10.2980/i1195-6860-12-3-316.1
Diamond, D. D., and C. D. True. 2008. Distribution of Juniperus woodlands in central Texas in relation to general abiotic site type. Pages 48-57 in O. W. Auken, editor. Western North American Juniperus communities. Springer, New York, New York, USA. https://doi.org/10.1007/978-0-387-34003-6_3
Ding, J., and D. J. Eldridge. 2019. Contrasting global effects of woody plant removal on ecosystem structure, function, and composition. Perspectives in Plant Ecology, Evolution, and Systematics 39:125460. https://doi.org/10.1016/j.ppees.2019.125460
Donovan, V. M., J. L. Burnett, C. H. Bielski, H. E. Birgé, R. Bevans, D. Twidwell, and C. R. Allen. 2018. Social-ecological landscape patterns predict woody encroachment from native tree plantings in a temperate grassland. Ecology and Evolution 8(19):9624-9632. https://doi.org/10.1002/ece3.4340
Driscoll, D. A., D. Armenteras, A. F. Bennett, L. Brotons, M. F. Clarke, T. S. Doherty, A. Haslem, L. T. Kelly, C. F. Sato, and H. Sitters. 2021. How fire interacts with habitat loss and fragmentation. Biological Reviews 96(3):976-998. https://doi.org/10.1111/brv.12687
Eidenshink, J., B. Schwind, K. Brewer, Z.-L. Zhu, B. Quayle, and S. Howard. 2007. A project for monitoring trends in burn severity. Fire Ecology 3:3-21. https://doi.org/10.4996/fireecology.0301003
Eldridge, D. J., L. Wang, and M. Ruiz‐Colmenero. 2015. Shrub encroachment alters the spatial patterns of infiltration. Ecohydrology 8(1):83-93. https://doi.org/10.1002/eco.1490
Ellis, E. C., and N. Ramankutty. 2008. Putting people in the map: anthropogenic biomes of the world. Frontiers in Ecology and the Environment 6(8):439-447. https://doi.org/10.1890/070062
Engle, D. M., B. R. Coppedge, and S. D. Fuhlendorf. 2008. From the dust bowl to the green glacier: human activity and environmental change in Great Plains grasslands. Pages 253-271 in O. W. Van Auken, editor. Western North American Juniperus communities, Ecological Studies volume 196, Springer, New York, New York, USA. https://doi.org/10.1007/978-0-387-34003-6_14
Fan, Y., J. Chen, G. Shirkey, R. John, S. R. Wu, H. Park, and C. Shao. 2016. Applications of structural equation modeling (SEM) in ecological studies: an updated review. Ecological Processes 5:1-12. https://doi.org/10.1186/s13717-016-0063-3
Fogarty, D. T., C. de Vries, C. Bielski, and D. Twidwell. 2021. Rapid re-encroachment by Juniperus virginiana after a single restoration treatment. Rangeland Ecology & Management 78:112-116. https://doi.org/10.1016/j.rama.2021.06.002
Fogarty, D. T., C. P. Roberts, D. R. Uden, V. M. Donovan, C. R. Allen, D. E. Naugle, M. O. Jones, B. W. Allred, and D. Twidwell. 2020. Woody plant encroachment and the sustainability of priority conservation areas. Sustainability 12:8321. https://doi.org/10.3390/su12208321
Fowler, N., and D. Dunlap. 1986. Grassland vegetation of the Eastern Edwards Plateau. American Midland Naturalist 115(1):146-155. https://doi.org/10.2307/2425844
Fuhlendorf, S. D., T. J. Hovick, R. D. Elmore, A. M. Tanner, D. M. Engle, and C. A. Davis. 2017. A hierarchical perspective to woody plant encroachment for conservation of prairie-chickens. Rangeland Ecology and Management 70(1):9-14. https://doi.org/10.1016/j.rama.2016.08.010
García Criado, M., I. H. Myers-Smith, A. D. Bjorkman, C. E. Lehmann, and N. Stevens. 2020. Woody plant encroachment intensifies under climate change across tundra and savanna biomes. Global Ecology and Biogeography 29(5):925-943. https://doi.org/10.1111/geb.13072
Gardner, R. 2009. Constructing a technological forest: nature, culture, and tree-planting in the Nebraska Sand Hills. Environmental History 14(2):275-297. https://doi.org/10.1093/envhis/14.2.275
Grant, T. A., E. Madden, and G. B. Berkey. 2004. Tree and shrub invasion in northern mixed-grass prairie: implications for breeding grassland birds. Wildlife Society Bulletin 32(3):807-818. https://doi.org/10.2193/0091-7648(2004)032[0807:TASIIN]2.0.CO;2
Gelbard, J. L., and J. Belnap. 2003. Roads as conduits for exotic plant invasions in a semiarid landscape. Conservation Biology 17(2):420-432. https://doi.org/10.1046/j.1523-1739.2003.01408.x
Harr, R. N., L. W. Morton, S. R. Rusk, D. M. Engle, J. R. Miller, and D. Debinski. 2014. Landowners’ perceptions of risk in grassland management: woody plant encroachment and prescribed fire. Ecology and Society 19(2):41. https://doi.org/10.5751/ES-06404-190241
Higgins, S. I., and S. Scheiter. 2012. Atmospheric CO₂ forces abrupt vegetation shifts locally, but not globally. Nature 488:209-212. https://doi.org/10.1038/nature11238
Higgins, S. I., C. M. Shackleton, and E. R. Robinson. 1999. Changes in woody community structure and composition under contrasting land use systems in a semi-arid savanna, South Africa. Journal of Biogeography 26(3):619-627. https://doi.org/10.1046/j.1365-2699.1999.t01-1-00317.x
Hoch, G., and J. Briggs. 1999. Expansion of Eastern Red Cedar in the Northern Flint Hills, Kansas. Pages 9-15 in Proceedings of the Sixteenth North American Prairie Conference (Kearney, 1998). University of Nebraska, Kearney, Nebraska, USA.
Hoekstra, J. M., T. M. Boucher, T. H. Ricketts, and C. Roberts. 2005. Confronting a biome crisis: global disparities of habitat loss and protection. Ecology Letters 8(1):23-29. https://doi.org/10.1111/j.1461-0248.2004.00686.x
Hoffman, J. K., R. P. Bixler, M. L. Treadwell, L. G. Coleman, T. W. McDaniel, and U. P. Kreuter. 2021. The impact of affective heuristics in decision-making regarding the implementation of prescribed fire on private rangelands in the Southern Great Plains, USA. Society & Natural Resources 34(5):1-18. https://doi.org/10.1080/08941920.2020.1864534
Hoy, J. 1989. Controlled pasture burning in the folk life of the Kansas Flint Hills. Great Plains Quarterly 9(4):231-238.
Kaur, R., O. Joshi, and R. E. Will. 2020. The ecological and economic determinants of eastern redcedar (Juniperus virginiana) encroachment in grassland and forested ecosystems: a case study from Oklahoma. Journal of Environmental Management 254:109815. https://doi.org/10.1016/j.jenvman.2019.109815
Koch, B., P. J. Edwards, W. U. Blanckenhorn, T. Walter, and G. Hofer. 2015. Shrub encroachment affects the diversity of plants, butterflies, and grasshoppers on two Swiss subalpine pastures. Arctic, Antarctic, and Alpine Research 47(2):345-357. https://doi.org/10.1657/AAAR0013-093
Kochy, M.,and S. D. Wilson. 2001. Nitrogen deposition and forest expansion in the northern Great Plains. Journal of Ecology 89(5):807-817. https://doi.org/10.1046/j.0022-0477.2001.00600.x
Kreuter, U. P., D. A. Stroman, C. L. Wonkka, J. Weir, A. A. Abney, and J. K. Hoffman. 2019. Landowner perceptions of legal liability for using prescribed fire in the Southern Plains, United States. Rangeland Ecology & Management 72:959-967. https://doi.org/10.1016/j.rama.2019.08.004
Jones, M. O., B. W. Allred, D. E. Naugle, J. D. Maestas, P. Donnelly, L. J. Metz, J. Karl, R. Smith, B. Bestelmeyer, and C. Boyd. 2018. Innovation in rangeland monitoring: annual, 30 m, plant functional type percent cover maps for US rangelands, 1984–2017. Ecosphere: Emerging Technologies 9(9):1-19. https://doi.org/10.1002/ecs2.2430
Lautenbach, J. M., R. T. Plumb, S. G. Robinson, C. A. Hagen, D. A. Haukos, and J. C. Pitman. 2017. Lesser Prairie-Chicken avoidance of trees in a grassland landscape. Rangeland Ecology & Management 70(1):78-86. https://doi.org/10.1016/j.rama.2016.07.008
Lockwood, J. L., P. Cassey, and T. Blackburn. 2005. The role of propagule pressure in explaining species invasions. Trends in Ecology & Evolution 20(5):223-228. https://doi.org/10.1016/j.tree.2005.02.004
Luvuno, L., R. Biggs, N. Stevens, and K. Esler. 2018. Woody encroachment as a social-ecological regime shift. Sustainability 10(7):2221. https://doi.org/10.3390/su10072221
Mohler, R. L., and D. G. Goodin. 2012. Mapping burned areas in the Flint Hills of Kansas and Oklahoma, 2000—2010. Great Plains Research 22(1):15-25.
Mosher, E. S., J. A. Silander, and A. M. Latimer. 2009. The role of land-use history in major invasions by woody plant species in the northeastern North American landscape. Biological Invasions 11(10):2317-2328. https://doi.org/10.1007/s10530-008-9418-8
Pickens, B. A., J. F. Marcus, J. P. Carpenter, S. Anderson, P. J. Taillie, and J. A. Collazo. 2017. The effect of urban growth on landscape-scale restoration for a fire-dependent songbird. Journal of Environmental Management 191:105-115. https://doi.org/10.1016/j.jenvman.2017.01.005
Picotte, J. J., K. Bhattarai, D. Howard, J. Lecker, J. Epting, B. Quayle, N. Benson, and K. Nelson. 2020. Changes to the monitoring trends in burn severity program mapping production procedures and data products. Fire Ecology 16:1-12. https://doi.org/10.1186/s42408-020-00076-y
Ratajczak, Z., J. M. Briggs, D. G. Goodin, L. Luo, R. L. Mohler, J. B. Nippert, and B. Obermeyer. 2016. Assessing the potential for transitions from tallgrass prairie to woodlands: are we operating beyond critical fire thresholds? Rangeland Ecology & Management 69(4):280-287. https://doi.org/10.1016/j.rama.2016.03.004
Ratajczak, Z., J. B. Nippert, and S. L. Collins. 2012. Woody encroachment decreases diversity across North American grasslands and savannas. Ecology 93(4):697-703. https://doi.org/10.1890/11-1199.1
Richardson, D. M., P. Williams, and R. J. Hobbs. 1994. Pine invasions in the Southern Hemisphere: determinants of spread and invadability. Journal of Biogeography 21(5):511-527. https://doi.org/10.2307/2845655
Roberts, C. P., D. R. Uden, C. R. Allen, and D. Twidwell. 2018. Doublethink and scale mismatch polarize policies for an invasive tree. PLoS ONE 13(3):e0189733. https://doi.org/10.1371/journal.pone.0189733
Roques, K. G., T. G. O’Connor, and A. R. Watkinson. 2001. Dynamics of shrub encroachment in an African savanna: relative influences of fire, herbivory, rainfall, and density dependence. Journal of Applied Ecology 38(2):268-280. https://doi.org/10.1046/j.1365-2664.2001.00567.x
Rosseel, Y. 2012. lavaan: an R Package for structural equation modeling. Journal of Statistical Software 48(2):1-36. https://doi.org/10.18637/jss.v048.i02
Roy, D. P., Y. Jin, P. E. Lewis, and C. O. Justice. 2005. Prototyping a global algorithm for systematic fire-affected area mapping using MODIS time series data. Remote Sensing of Environment 97:137-162. https://doi.org/10.1016/j.rse.2005.04.007
Sankaran, M., N. P. Hanan, R. J. Scholes, J. Ratnam, D. J. Augustine, B. S. Cade, J. Gignoux, S. I. Higgins, X. Le Roux, and F. Ludwig. 2005. Determinants of woody cover in African savannas. Nature 438:846-849. https://doi.org/10.1038/nature04070
Scholes, R., and S. Archer. 1997. Tree-grass interactions in savannas. Annual Review of Ecology and Systematics 28:517-544. https://doi.org/10.1146/annurev.ecolsys.28.1.517
Scholtz, R., S. D. Fuhlendorf, and S. Archer. 2018b. Climate-fire interactions constrain potential woody plant cover and stature in North American Great Plains grasslands. Global Ecology and Biogeography 27(8):936-945.
Scholtz, R., S. D. Fuhlendorf, D. R. Uden, B. W. Allred, M. O. Jones, D. E. Naugle, and D. E. Twidwell. 2021. Challenges of brush management treatment effectiveness in Southern Great Plains, United States. Rangeland Ecology & Management 77:57-65. https://doi.org/10.1016/j.rama.2021.03.007
Scholtz, R., J. Polo, E. Tanner, and S. Fuhlendorf. 2018a. Grassland fragmentation and its influence on woody plant cover in the Southern Great Plains, USA. Landscape Ecology 33:1785-1797. https://doi.org/10.1007/s10980-018-0702-4
Sharp, E. A., P. G. Spooner, J. Millar, and S. V. Briggs. 2012. Can’t see the grass for the trees? Community values and perceptions of tree and shrub encroachment in south-eastern Australia. Landscape and Urban Planning 104(2):260-269. https://doi.org/10.1016/j.landurbplan.2011.11.009
Shigesada, N., and K. Kawasaki. 1997. Biological invasions: theory and practice. Oxford University Press, Oxford, UK.
Stevens, N., C. E. Lehmann, B. P. Murphy, and G. Durigan. 2017. Savanna woody encroachment is widespread across three continents. Global Change Biology 23(1):235-244. https://doi.org/10.1111/gcb.13409
Stroman, D. A., and U. P. Kreuter. 2014. Perpetual conservation easements and landowners: evaluating easement knowledge, satisfaction, and partner organization relationships. Journal of Environmental Management 146:284-291. https://doi.org/10.1016/j.jenvman.2014.08.007
Stroman, D. A., U. P. Kreuter, and C. L. Wonkka. 2020. Landowner perceptions of woody plants and prescribed fire in the Southern Plains, USA. PLoS ONE 15(9):e0238688. https://doi.org/10.1371/journal.pone.0238688
Sullins, D. S., D. A. Haukos, J. M. Lautenbach, J. D. Lautenbach, S. G. Robinson, M. B. Rice, B. K. Sandercock, J. D. Kraft, R. T. Plumb, and J. H. Reitz. 2019. Strategic conservation for Lesser Prairie-Chickens among landscapes of varying anthropogenic influence. Biological Conservation 238:108213. https://doi.org/10.1016/j.biocon.2019.108213
Toledo, D., U. P. Kreuter, M. G. Sorice, and C. A. Taylor, Jr. 2014. The role of prescribed burn associations in the application of prescribed fires in rangeland ecosystems. Journal of Environmental Management 132:323-328. https://doi.org/10.1016/j.jenvman.2013.11.014
Toledo, D., M. G. Sorice, and U. P. Kreuter. 2013. Social and ecological factors influencing attitudes toward the application of high-intensity prescribed burns to restore fire adapted grassland ecosystems. Ecology and Society 18(4):9. https://doi.org/10.5751/ES-05820-180409
Twidwell, D., W. E. Rogers, S. D. Fuhlendorf, C. L. Wonkka, D. M. Engle, J. R. Weir, U. P. Kreuter, and C. A. Taylor. 2013. The rising Great Plains fire campaign: citizens’ response to woody plant encroachment. Frontiers in Ecology and the Environment 11(s1):e64-e71.
Twidwell, D., W. E. Rogers, C. L. Wonkka, C. A. Taylor Jr, and U. P. Kreuter. 2016b. Extreme prescribed fire during drought reduces survival and density of woody resprouters. Journal of Applied Ecology 53(5):1585-1596. https://doi.org/10.1111/1365-2664.12674
Twidwell, D., A. S. West, W. B. Hiatt, A. L. Ramirez, J. T. Winter, D. M. Engle, S. D. Fuhlendorf, and J. Carlson. 2016a. Plant invasions or fire policy: which has altered fire behavior more in tallgrass prairie? Ecosystems 19:356-368. https://doi.org/10.1007/s10021-015-9937-y
Twidwell, D., C. L. Wonkka, H.-H. Wang, W. E. Grant, C. R. Allen, S. D. Fuhlendorf, A. S. Garmestani, D. G. Angeler, C. A. Taylor Jr, and U. P. Kreuter. 2019. Coerced resilience in fire management. Journal of Environmental Management 240:368-373. https://doi.org/10.1016/j.jenvman.2019.02.073
Van Auken, O. W. 2000. Shrub invasions of North American semiarid grasslands. Annual Review of Ecology and Systematics 31:197-215. https://doi.org/10.1146/annurev.ecolsys.31.1.197
Van Auken, O. W. 2009. Causes and consequences of woody plant encroachment into western North American grasslands. Journal of Environmental Management 90(10):2931-2942. https://doi.org/10.1016/j.jenvman.2009.04.023
Venter, Z. S., M. D. Cramer, and H.-J. Hawkins. 2018. Drivers of woody plant encroachment over Africa. Nature Communications 9:2272. https://doi.org/10.1038/s41467-018-04616-8
Wells, P. V. 1965. Scarp woodlands, transported grassland soils, and concept of grassland climate in the Great Plains Region. Science 148(3667):246-249.
Wilcox, B. P., A. Birt, S. R. Archer, S. D. Fuhlendorf, U. P. Kreuter, M. G. Sorice, W. J. van Leeuwen, and C. B. Zou. 2018. Viewing woody-plant encroachment through a social-ecological lens. BioScience 68(9):691-705. https://doi.org/10.1093/biosci/biy051
Woods, N. N., B. L. Dows, E. B. Goldstein, L. J. Moore, D. R. Young, and J. C. Zinnert. 2019. Interaction of seed dispersal and environmental filtering affects woody encroachment patterns in coastal grassland. Ecosphere 10(7):e02818. https://doi.org/10.1002/ecs2.2818
Yoder, J., D. Engle, and S. Fuhlendorf. 2004. Liability, incentives, and prescribed fire for ecosystem management. Frontiers in Ecology and the Environment 2(7):361-366. https://doi.org/10.1890/1540-9295(2004)002[0361:LIAPFF]2.0.CO;2
Yokomizo, H., H. P. Possingham, M. B. Thomas, and Y. M. Buckley. 2009. Managing the impact of invasive species: the value of knowing the density-impact curve. Ecological Applications 19(2):376-386. https://doi.org/10.1890/08-0442.1
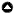
Fig. 1

Fig. 1. Three study ecoregions, which include the Flint Hills, Rolling Red Hills, and Edwards Plateau (regions outlined in bold dark lines) within the Southern Great Plains ecoregion (grey) of North America, USA.
Fig. 2

Fig. 2. Estimated rate of change in woody plant cover per year at the county level (A) and the estimated percent change (B) between 2000 and 2019 in woody plant cover for regions within the Edwards Plateau, Rolling Red Hills, and Flint Hills ecoregions in the Southern Great Plains, USA.
Fig. 3

Fig. 3. Path diagram showing the relationships between anthropogenic and biophysical variables with each other and rates of woody plant encroachment in the Southern Great Plains, USA. Boxes overlaid on significant paths show the standardized parameter estimate and the estimate’s p-value. Estimates and p-values are not shown for nonsignificant pathways (greyed lines).
Table 1
Table 1. Anthropogenic and biophysical variables used to assess rates of county level woody plant encroachment in the Edwards Plateau, Flint Hills, and Rolling Red Hills ecoregions in the Southern Great Plains, USA, from 2000 to 2019. All variables, average percent area burned, represent conditions at the beginning of the study period (2000–2001) as we were interested in how initial conditions would influence subsequent rates of encroachment. The mean and standard deviation (parentheses) of the county level data are shown for each ecoregion.
| Edwards Plateau | Flint Hills | Rolling Red Hill | Data Source | |
| Biophysical Variables | ||||
| Average percent area burned | 0.17 (0.35) | 4.73 (4.58) | 0.47 (0.81) | Monitoring Trends in Burn Severity Database |
| Percent Cover Trees (%) | 24.99 (11.09) | 8.23 (3.68) | 7.13 (3.32) | Rangeland Analysis Platform |
| Mean Annual Precipitation (mm) |
682.59 (142.36) | 905.69 (74.87) | 672.26 (51.85) | PRISM Climate Data |
| Anthropogenic Variables | ||||
| Population Density (people per km²) | 39 (107.96) | 17.15 (27.73) | 4.71 (3.42) | 2000 census |
| Percent Cover Agriculture (%) | 4.9 (7.17) | 32.02 (15.93) | 27.86 (17.15) | National Land Cover Dataset |
| Percent Cover Developed (%) | 5.44 (6.69) | 5.36 (3.07) | 4.63 (0.7) | National Land Cover Dataset |
| Road Density (roads per km²) | 0.68 (0.85) | 1.39 (0.38) | 0.6 (0.16) | National Tiger Roads GIS |
Table 2
Table 2. Hypotheses describing the direct and indirect relationships between rates of woody plant encroachment and anthropogenic and biophysical variables in three ecoregions of the Southern Great Plains, USA, from 2000 to 2019. In some cases, two or more hypotheses offer alternative explanations for the link between two variables and may even have contrasting predictions (one hypothesis may predict a positive effect while another predicts a negative effect). Hypotheses were used to determine the structural equation model used in subsequent analysis.
| Direct effects | Citation | |
| Hypothesis 1 | Counties that experienced more fires will have lower rates of woody plant encroachment, because fires will limit the establishment of new trees. | Sholes and Archer 1997, Briggs et al. 2005, Archer et al. 2017 |
| Hypothesis 2 | Counties with higher initial tree cover will have higher rates of woody plant encroachment, because these already established trees can act as propagule sources for new trees. | Donovan et al. 2018, Woods et al. 2019 |
| Hypothesis 3 | Fragmentation of grasslands from anthropogenic sources such as roads, development, agriculture, or trees will alter rates of encroachment either by limiting areas where trees can spread or because of alternative land uses that limits management for trees. | Gelbard et al. 2003, Mosher et al. 2009, Archer et al. 2017, Scholtz et al. 2018b |
| Hypothesis 4 | Higher annual precipitation will allow for higher rates of encroachment, as woody cover potential has been shown to be related to precipitation patterns. | Sankaran et al. 2005, Scholtz et al. 2018a |
Indirect effects |
||
| Hypothesis 5 | Counties with higher levels of fragmentation of grassland will have reduced fire activity leading to greater woody plant encroachment. This may be due to limited spread of fires, or because of a greater diversity of land use practices that may not facilitate or allow for prescribed burning. | Scholtz et al. 2018b, Donovan et al. 2020, Driscoll et al. 2021 |
| Hypothesis 6 | Counties with higher levels of fragmentation or population density will have higher initial tree cover because trees are intentionally planted near human structures, or because variable/inconsistent management practices have allowed trees to become established around human structures. This may alter encroachment rates through either increased propagule sources (higher encroachment) or increased competition (lower encroachment). | Hoch and Briggs 1999, Gelbard et al. 2003, Gardner 2009, Scholtz et al. 2018b |
| Hypothesis 7 | Counties with higher population densities will have fewer fires resulting in greater woody plant encroachment. This can be the result of increased risk of liability for managers using prescribed fire, local regulations regarding smoke management, or increased use of land use practices that may not facilitate or allow for prescribed burning. | Hoch and Briggs 1999, Berg et al. 2015 |
Table 3
Table 3. Area covered by trees (percent area covered by trees in parenthesis) in the Edwards Plateau, Flint Hills, and Rolling Red Hills ecoregions in the Southern Great Plains, USA, at the start of the study period (2000) and end of the study period (2019). Additionally, we present the percent change in cover of trees in each ecoregion from 2000 to 2019.
| Region | Initial tree cover in km² | Final tree cover in km² | Percent change |
| Flint Hills | 180770 (6.4%) | 265002 (9.4%) | 46.59% |
| Edwards Plateau | 2002122 (26.7%) | 2375052.5 (31.68) | 18.62% |
| Rolling Red Hills | 101763.8 (7.7%) | 141659.9 (10.7%) | 39.20% |