






The following is the established format for referencing this article:
Herse, M. R., P. O'B. Lyver, A. M. Gormley, N. J. Scott, A. R. McIntosh, D. Fletcher, and J. Tylianakis. 2022. A demographic model to support customary management of a culturally important waterfowl species. Ecology and Society 27(3):14.ABSTRACT
Scientific support invited by Indigenous peoples and local communities (IPLC) to assist with customary environmental management can improve conservation and community livelihoods. For example, demographic models can help to understand how alternative wildlife management strategies affect population dynamics and harvest sustainability. We developed a demographic model to assist Ngāi Tahu, the southern-most Māori tribe in Aotearoa/New Zealand, in customary management of a culturally important population of Black Swans (kakī anau, Cygnus atratus). We used recent demographic data, including results of an experimental egg harvest study, to inform tangata tiaki (Ngāi Tahu environmental guardians) about how customary egg harvest and background pressure from sport hunting of swans aged ≥ 1 year differentially affect population growth. We also assessed how sport hunting of swans affects the sustainability of customary egg harvest. Estimated population growth (1.018 or presently growing 1.8% annually; 95% CI: 0.808–1.241) was most sensitive to changes in adult and subadult survival, followed by juvenile (first-year) survival, breeding propensity, and nest hatching success. Uncertainty in population growth was almost entirely attributable to uncertainty in swan survival rates after hatching. Sustainable population-level rates of egg harvest varied from none to more than half of all eggs, depending on small changes in adult and subadult survival. Population sensitivity to adult and subadult survival suggests that limiting and monitoring their mortality are crucial to population and egg harvest sustainability, whereas contemporary government-mandated species management, through Fish and Game New Zealand, allows adult and subadult mortality from sport hunting, with little record of offtake. Recognizing the rights and interests of Ngāi Tahu, and monitoring swan mortality more closely, could improve Ngāi Tahu abilities to practice customary harvest, enhance population and environmental monitoring, and, when appropriate, control swan numbers in a culturally appropriate and less wasteful way. The model we present could aid decision making and communication between Ngāi Tahu and New Zealand’s Crown government within a potential future co-management arrangement. Demographic models can be useful tools for supporting customary environmental management, but developing, maintaining, and implementing these tools requires support for adaptive policies and management arrangements that recognize IPLC rights to the environment and decision making.INTRODUCTION
Contemporary environmental management systems are struggling to curb global declines in biodiversity and ecosystem health (Díaz et al. 2019, IPBES 2019a). One limiting factor for management is the ability to continually monitor biodiversity and ecosystems at fine resolutions across large areas, and to implement adaptive conservation measures locally (Cumming et al. 2006, Guerrero et al. 2013). To help overcome this limitation, and to re-claim rights and responsibilities for the environment more generally, many Indigenous peoples and local communities (IPLC or “ethnic groups who are descended from and identify with the original inhabitants of a given region, in contrast to groups that have settled, occupied or colonized the area more recently”; IPBES 2019b) are vocal and active in empowering their customary environmental management systems (Ens et al. 2012, Witter et al. 2015, Lyver and Tylianakis 2017). Within many customary management systems, IPLC continually engage with and monitor their environments, while over time accumulating long-term experiential knowledge of biodiversity and ecosystems at fine resolutions (Gadgil et al. 1993, Moller et al. 2004, Berkes 2009). Customary resource use and engagement with the environment underpin many aspects of well-being for IPLC, including livelihoods, cultural identities, and community kinship, and these relationships can motivate sustainable practices and enforcement of local conservation (Cinner and Aswani 2007, Lyver et al. 2008, Halim et al. 2012, Sheil et al. 2015, Caillon et al. 2017). Thus, supporting IPLC in the customary management of their lands and resources could be both a practical and just way to enhance environmental monitoring and protection (Sobrevila 2008, Artelle et al. 2019, Herse et al. 2020, Dawson et al. 2021, Reyes-García et al. 2022).
In many places, the legal authority of IPLC to decide whether resource use can occur, what uses and strategies are appropriate, and who should be allowed access to use, are either missing or neglected (Kinan and Dalzell 2005, Goldman 2011, Ruru et al. 2017, Snook et al. 2020). In such cases, management decisions by non-IPLC authorities are often biased toward competing interests of recreational or commercial harvest (Ban et al. 2017), alternative land uses, e.g., mining (Parlee et al. 2018), or preservationist strategies, e.g., protected areas, that prioritize separation between IPLC and their environment (Brockington and Igoe 2006, Solomon 2014) and do not value customary harvests (Hunn et al. 2003, Tyrrell 2008, Lyver et al. 2019a). Environmental scientists have increasingly used demographic models of animal populations to inform decision making about customary resource use and management (Brook et al. 2006, Zador et al. 2006, Bradshaw and Brook 2007, Chaloupka and Balazs 2007, Jones et al. 2015, McKechnie et al. 2020). For example, Chaloupka and Balazs (2007) used a demographic model to inform public policy debates in Hawaiʻi, USA, about the sustainability of re-initiating customary harvests of green sea turtles (Chelonia mydas), which were prohibited following overexploitation by commercial harvest and subsequent listing of the green sea turtle as an “endangered species.” In Australia, Bradshaw and Brook (2007) used demographic models to assess strategies for balancing customary use of exotic, endangered banteng (Bos javanicus) by Aboriginal communities, and offtake of recreational and commercial harvest. However, IPLC voices and interpretations of scientific evidence are largely absent from such studies if they are not initiated by IPLC or do not include IPLC representation. Likewise, when IPLC wish to engage in customary management and welcome scientific support, collecting, interpreting, and disseminating data in partnership with IPLC is important to ensure that their rights, values, and priorities are fully recognized (Ens et al. 2012, Rayne et al. 2020, Dawson et al. 2021).
Here we present a demographic model developed in partnership with Ngāi Tahu, the southern-most tribe of Māori in Aotearoa/New Zealand, to support customary management of Black Swans (kakī anau, Cygnus atratus) at a 200-km² coastal lagoon and key Black Swan breeding area, Te Waihora/Lake Ellesmere (Fig. 1). Harvests of Black Swans and their eggs have been important components of customary management and food gathering at Te Waihora for generations, but restricted since New Zealand’s colonial government seized control of local species management in the early 20th century CE (Lamb 1964, Pauling and Arnold 2008, Herse et al. 2021). Current law in New Zealand classifies Black Swans as “gamebirds” that can be hunted for sport by gamebird license holders (New Zealand Government 1953), yet prohibits customary harvest without special permission (Herse et al. 2020, 2021). Contemporary government-mandated gamebird managers use daily bag limits (quotas), set annually, to maintain swan numbers in the recreational interests of sport hunters (New Zealand Government 1987), and in the past have permitted culls by local farmers (European New Zealanders) to limit grazing of pasture and potential economic loss (Williams 1979). The extent of pressure from sport hunting and past culling is poorly known because the contemporary gamebird management body, Fish and Game New Zealand (FGNZ), does not require licensed sport hunters or landowners to report their offtake, and periodic regional phone surveys by FGNZ of offtake trends are not site specific (Herse et al. 2020).
Ngāi Tahu is interested in managing the swan population at least partly through self-authorized customary egg harvests, without oversight or special permission (Herse et al. 2020, 2021). The right of Ngāi Tahu to self-authorize harvest and management of swans is guaranteed within New Zealand’s founding document between tangata whenua (original people of the land) and the Crown government, the Treaty of Waitangi 1840, whereby “the Queen of England agrees to protect the chiefs, the subtribes and all the people of New Zealand in the unqualified exercise of their chieftainship over their lands, villages and all their treasures” (Kawharu 1989:321). Similarly, Ngāi Tahu rights to “determine and develop priorities and strategies for the development or use of their lands or territories and other resources” are protected under the United Nations Declaration on the Rights of Indigenous Peoples (United Nations General Assembly 2007:23). However, these rights have not been realized or enacted under current gamebird management regulations. In addition to FGNZ-permitted sport hunting and culling, damage to aquatic plants by a massive storm in 1968 (the “Wahine” storm), coupled with extensive agricultural runoff over the past half century, has degraded Te Waihora, depleted swans’ macrophyte food supply (Lineham 1983, Gerbeaux 1989, Hearnshaw and Hughey 2010), and reduced swan numbers by approximately 90% (Williams 1979, Ford et al. 2017). Thus, it is unclear whether the current swan population can sustain potential customary egg harvest or any of the ongoing sport hunting, and how such pressures on swan fertility (hatching success) and survivorship, respectively, affect the population.
Our goal was to provide Ngāi Tahu with up-to-date information on the Te Waihora swan population with which to enter discussions with FGNZ and New Zealand’s Crown government, about potential strategies for re-initiating customary egg harvests. To that end, we started by using recent demographic data and an age-classified matrix population model to (a) estimate the current annual population growth rate or change in population size (λ; Caswell 2001). We then used the model to assess (b) how changes in swan fertility and age-specific survivorship differentially affected λ, and (c) how current levels of uncertainty in these demographic rates differentially contributed to uncertainty in λ. To understand the egg-harvest equivalence of sport hunting, we estimated (d) the expected lifelong output of both freshly laid eggs and hatched eggs (i.e., measures of reproductive value; Caswell 2001) by individual swans aged ≥ 1 year, which are collectively most susceptible to being shot (Williams 1977). Finally, we used results of a recent experimental egg harvest study (Herse et al. 2021) to (e) assess how offtake from sport hunting affects the sustainability of potential customary egg harvest by Ngāi Tahu. Based on the model results, we explore potential pathways for incorporating customary egg harvest into local species management. We end by outlining potential ways of improving support for customary management, which could enhance environmental monitoring and protection (Ens et al. 2016, Herse et al. 2020, Dawson et al. 2021, Lyver et al. 2021).
METHODS
Study species and population
The modern Black Swan (C. atratus) originates from Australia and is congeneric with the endemic New Zealand Black Swan (poūwa, C. sumnerensis), which became extinct on New Zealand’s mainland around the 15th century, after Māori arrival and prior to European arrival (Rawlence et al. 2017). Europeans reported C. atratus as absent in New Zealand during the 19th century and introduced several pairs from Australia in the 1860s (Lamb 1964). However, rapid population growth following the introductions, and recent DNA evidence, suggest that waves of C. atratus dispersal from Australia before and during the 19th century, and/or genetic rescue of an extant population through admixing with introduced individuals, may have occurred (Rawlence et al. 2017, Montano et al. 2018a). Consequently, scientists have debated whether modern Black Swans are native to New Zealand (Montano et al. 2018b, Rawlence et al. 2018). Some Ngāi Tahi tribal members consider Black Swans native to New Zealand (Bataille 2020), and the species is recognized as such within the Te Waihora Joint Management Plan between Ngāi Tahu and the Minister of Conservation (Te Rūnanga o Ngāi Tahu and Department of Conservation 2005). More importantly, Black Swans are highly valued by some Ngāi Tahu people as key elements of cultural practices such as kaitiakitanga (environmental guardianship) and mahinga kai (“customary gathering of food and natural materials and the places where those resources are gathered”; New Zealand Government 1998:102, Bataille 2020, Bataille et al. 2021), which are important to Māori cultural integrity (Kawharu 2000), knowledge production (Lyver et al. 2019b), and conservation (Wehi and Lord 2017). The species is also valued by some gamebird license holders and other members of the public, whereas some farmers consider swans to be pests because they sometimes graze on pasture (Bataille et al. 2020, 2021).
Black Swans (hereafter “swans”) mostly breed in colonies at shallow lakes, lagoons, and estuaries (Williams 1981, 2013a). Te Waihora contains the largest population of swans on New Zealand’s South Island, with estimates of population size varying from ~5000 to 10,000 individuals during the past two decades (Ford et al. 2017, Herse et al. 2020). Similar to other swan species globally, most Black Swans at Te Waihora live for fewer than 10 years, and less than half of adults attempt nesting each year (Williams 1979, 1981, 2013, Bart et al. 1991). Black Swans are typically monogamous and breeding at Te Waihora is generally synchronous, beginning in August to September as winter water levels decline, and decreasing in October after most nests hatch (Williams 1981, 2013a, Herse et al. 2021). Mean clutch size is 5–6 eggs (Williams 2013a, Herse et al. 2021). A second bout of breeding, which comprises up to approximately one-third of all nesting during a given breeding season, sometimes occurs during October–December (Donald Brown, Ngāi Tahu, Christchurch, New Zealand, 19 June 2018, personal communication; Herse et al. 2021). In a previous experimental egg harvest study, Herse et al. (2021) found that following the complete removal of first clutches of eggs, swans tended to replace a portion of those eggs, such that the average nest-level hatching success among harvested nests was 2.61 cygnets (cf. 5.68 cygnets per unharvested nest). Although Herse et al. (2021) could not determine whether re-nesting following clutch failure was by original nest occupants versus other swans using newly available nests, individual pairs have been documented re-nesting following clutch failure (Williams 1979), indicating that at least some of the second bout of breeding is due to re-nesting. Pressures on nest hatching success in our study area include nest depredation, flooding, and infertility, but hatching success is nevertheless generally high at the largest nesting colonies (Adams 1971, Meis 1988, Herse et al. 2021). Juvenile swans (cygnets) reach independence ~3–5 months after hatching and sexual maturity at 2–4 years of age (Williams 2013a).
In addition to food availability, current pressures on swan survivorship include sport hunting (Williams 1979, Herse et al. 2020). Gamebird managers set bag limits based on trends in swan numbers and offtake rates estimated from periodic aerial surveys and phone surveys, respectively, and aggregated at scales of hunting regions or subregions over which individual bag limits apply (Herse et al. 2020). During recent years, gamebird license holders within the ~2,500-km² hunting subregion containing Te Waihora have each been allowed to shoot 5 swans per day during most of May–July, and 20 swans per day during most of August–September (Herse et al. 2020). However, current rates of gamebird shooting and their impacts on the swan population at Te Waihora are poorly understood for several reasons. New Zealand law requires private landowners to adhere to local gamebird bag limits but does not require landowners to obtain licenses to shoot swans on their property (New Zealand Government 1953), or to report their offtake. Similarly, FGNZ does not require gamebird license holders to report their offtake. In addition, regional surveys of offtake are not site specific and include areas outside of the Te Waihora subregion (Herse et al. 2020).
Matrix population model
We developed an age-classified matrix population model (Leslie 1945, Caswell 2001) to describe swan population dynamics. The model is defined by
 |
(1) |
where A is a projection matrix including age-specific, per-capita estimates of swan fertility and survival rates, and nt is a vector of numbers of individuals per age class during time t (Caswell 2001). Time units span one year beginning when eggs hatch and ending immediately prior to hatching during the following nesting season (“post-breeding census”; Caswell 2001). We assumed the population was closed to immigration and emigration because swan dispersal among waterbodies within the region is infrequent (Williams 1977; Herse, Lyver, Scott, Tylianakis, and Ngāi Tahu, unpublished data), and local species management cannot control movements of the population’s small proportion of dispersive individuals.
Considering age at first reproduction is 2–4 years (Williams 2013a), it would be reasonable to develop a stage-classified model based on hatchling, sexually immature subadult, and adult stage classes. However, because we could not distinguish subadults from non-breeding adults during population censuses, we could not estimate the proportion of adults attempting breeding per year; we could only estimate this proportion for subadults and adults combined. In addition, we lacked data on the current age distribution of swans at Te Waihora, and did not know if or how ongoing mortality from sport hunting varied across annual age classes. Thus, we based the model on a simplified life cycle with hatch-year (HY, age < 1 year since hatching) and after-hatch-year (AHY, age ≥ 1 year since hatching) age classes (Fig. 2). The fertility term in this model includes the proportion of AHY swans, i.e., subadults plus adults, that attempt breeding each year, which we could estimate. Demographic rates of finer resolution age or stage classes could be parameterized later with additional data if they become available.
The model can be written in matrix form as
 |
(2) |
where NHY,t and NAHY,t are the number of HY and AHY swans, respectively, during time (year) t; FAHY is annual fertility or number of eggs hatched per AHY swan; SHY is the probability of a swan surviving the first year after hatching (i.e., the “hatch year”); and SAHY is annual survival probability for AHY swans. Fertility of HY swans (top left element of matrix in Equation 2) is 0 because HY swans do not breed (Williams 2013). Modeled demographic transition rates (matrix elements) are time-invariant, but could be updated annually with new data.
Factors that affect annual fertility of AHY swans include propensity to attempt breeding, nest hatching success, and mortality of AHY swans (Williams 1979, 1981, Herse et al. 2021). Hatching success differs between unharvested and harvested nests because, in recent years, swans tend to replace only a subset of harvested eggs (Herse et al. 2021). Thus, we estimated FAHY using the expression:
 |
(3) |
where f0 is fertility per unharvested nest, fa is fertility per harvested nest, h is the proportion of total nests harvested, and b is the proportion of AHY swans that attempt breeding, as noted above. Multiplying fertility per nest by 0.5 gives fertility per breeding swan, because breeding is typically monogamous (Williams 2013a). The model assumes that during fixed-output egg harvests, harvesters remove entire clutches of eggs from harvested nests (as opposed to removing fewer eggs per nest from a larger number of nests), because this was the strategy found to have the least impact on population-level hatching success during a recent harvest experiment (Herse et al. 2021). The SAHY term in Equation 3 accounts for some swans not surviving the current year to attempt breeding in year t + 1.
The limited availability of macrophyte food at Te Waihora could prevent the swan population from increasing above a certain size, regardless of pressure from sport hunting or customary harvest (Williams 1979, Herse et al. 2021). However, we did not include density dependence in the model because we lacked information to define a swan population carrying capacity for Te Waihora.
Estimation of demographic parameters
We estimated demographic parameters and associated variances in three steps using a parametric bootstrap approach that included empirical data and indirect methods (Fig. 3). First, we used recent demographic data to define initial bootstrap sampling distributions for estimates of λ, SAHY, and underlying parameters of FAHY (Equation 3, Table 1, Fig. 3). Second, we used random sampling from the initial distributions to back-calculate, and hence generate a bootstrap distribution for, estimates of SHY, which we lacked data to confidently estimate directly (Table 1, Fig. 3). Back-calculations of SHY incorporated information on survival of swan hatchlings and resulted in adjusted bootstrap distributions for SAHY and underlying parameters of FAHY (Table 1, Fig. 3). Finally, we used random sampling from the resulting bootstrap distributions to parameterize the matrix model and generate bootstrap distributions for a suite of demographic parameters for the Te Waihora population (Table 2, Fig. 3). All bootstrap distributions were based on n = 10,000 samples. We provide more detail on each of these steps below and in Appendix 1.
To define initial sampling distributions used for back-calculating SHY, we estimated NHY and NAHY based on mean counts of swans made by two independent observers during a post-breeding aerial census in December, 2018 (Fig. 3, see Appendix 1). We estimated f0 and fa using data from observations of unharvested and harvested nests, respectively, during an experimental egg harvest at Te Waihora during the peak nesting period (August–September) in 2018 (Fig. 3, Fig. A1.1, see Appendix 1; Herse et al. 2021). Thus, the model assumes that all breeding during a given year occurs during peak nesting season, which we visually estimated to include ~88% of all nests at Te Waihora in 2018 (see Appendix 1). However, the model accounts for contributions of late-season nesting to estimated population growth rate,  , indirectly through back-calculations of SHY (detailed below), which incorporated information on NHY and total population size, including cygnets hatched from late season nests. We estimated b as the proportion of AHY swans that attempted breeding during the peak nesting period at Te Waihora in 2018, as determined by aerial censuses of nests (assuming each nest represented two breeding swans; Williams 2013a) and AHY swans (Fig. 3, Fig. A1.2, see Appendix 1). The estimate of b in 2018 (0.30) was similar to estimates from previous studies (0.16–0.34; Williams 1981). We estimated SAHY based on known fates of 17 GPS-marked AHY swans we monitored at Te Waihora during 2019–2020 (Fig. 3, see Appendix 1), which had an estimated mean survival rate similar to those estimated from previous long-term band-recovery studies (Fig. 3, Fig. A1.3; Williams 1979, Barker and Buchanan 1993).
, indirectly through back-calculations of SHY (detailed below), which incorporated information on NHY and total population size, including cygnets hatched from late season nests. We estimated b as the proportion of AHY swans that attempted breeding during the peak nesting period at Te Waihora in 2018, as determined by aerial censuses of nests (assuming each nest represented two breeding swans; Williams 2013a) and AHY swans (Fig. 3, Fig. A1.2, see Appendix 1). The estimate of b in 2018 (0.30) was similar to estimates from previous studies (0.16–0.34; Williams 1981). We estimated SAHY based on known fates of 17 GPS-marked AHY swans we monitored at Te Waihora during 2019–2020 (Fig. 3, see Appendix 1), which had an estimated mean survival rate similar to those estimated from previous long-term band-recovery studies (Fig. 3, Fig. A1.3; Williams 1979, Barker and Buchanan 1993).
We used random samples from the initial bootstrap distributions described above to back-calculate SHY. Because population growth rate, λ, the dominant eigenvalue of matrix A, can be solved by the characteristic equation:
 |
(4) |
where I is a 2 × 2 identity matrix (Caswell 2001), we rearranged Equation 4 and estimated SHY using the expression:
 |
(5) |
where λ was estimated based on the annual trend in swan population size at Te Waihora during 2017–2019, as determined by aerial censuses (see Appendix 1). Specifically, we estimated λ in Equation 5 as the exponent of the slope of the linear regression line relating log population size to year (Fig. 3, Fig. A1.4; see Appendix 1; Peery et al. 2006). We applied several constraints to Equation 5 to ensure that ŜHY was > 0 and less than the estimated survival rate of n = 50 marked swan hatchlings over their first 40 days of life at Te Waihora in 2018 (Fig. 3; see Appendix 1). If constraints were not met during a draw of samples, we discarded samples of all parameters in that draw and sampled again. The mean of the resulting bootstrap distribution for SHY (Table 1, Fig. 3) was congruent with estimates extrapolated from a survival analysis of the swan hatchling data and estimates from previous studies (Fig. A1.5, see Appendix 1; Williams 1979, Meis 1988). The application of constraints to back-calculations of SHY resulted in adjusted bootstrap distributions of parameters used in back-calculations (Table 2, Fig. 3).
We used random samples from bootstrap distributions described above to parameterize the matrix model (Table 1, Fig. 3). We used functions available in the R “popbio” package (Stubben et al. 2020, R Core Team 2021) to estimate a suite of demographic parameters for the swan population, assuming no egg harvest pressure (h = 0). We estimated annual λ as the dominant eigenvalue of the matrix A (Fig. 3), as described above (Caswell 2001). To determine how changes in SHY and SAHY, and lower-level rates of FAHY (f0 and b), differentially affected λ, we calculated sensitivity (sx) and elasticity (ex) of to absolute and proportional changes, respectively, in each underlying demographic rate x (see Appendix 1; Caswell 2001). Elasticity can be interpreted as a measure of the relative importance of a given demographic rate for determining λ (Caswell 2001). Both sensitivity and elasticity are useful and simply measure effects of changes in demographic rates at different scales (i.e., absolute versus proportional); measuring both ensured that we would not overlook an effect that is apparent on only one scale (Caswell 2001). To assess how current levels of uncertainty in demographic rates differentially contributed to uncertainty in λ, we used variances of bootstrap distributions of sx and ex to calculate the proportional contribution of each demographic rate to the total variance in λ (see Appendix 1; Caswell 2001). To determine the egg-harvest equivalence of removing one AHY swan from the population, we estimated the life expectancy and reproductive value (i.e., expected per-capita lifelong reproductive output, rHY, or number of HY swans produced; Caswell 2001) of AHY swans, and then estimated the expected per-capita lifelong number of fresh eggs laid as
 |
(6) |
where Segg is the estimated mean survival (hatching) probability of an individual egg from an unharvested nest (Herse et al. 2021).
Sustainability of mixed management
To assess how offtake from sport hunting of AHY swans affects the sustainability of potential customary egg harvests, we used the matrix model to estimate λ based on varying rates of SAHY and h individually and in combination. Te Waihora-specific information on sport hunting of swans does not exist, as described above (Herse et al. 2020), and our estimate of SAHY accounts for risk from this pressure. Thus, we considered a range of SAHY covering the model estimated mean SAHY ± 0.1 (in increments of 0.02), which was a wide enough range to detect both positive and negative across all possible rates of h. We considered the full range of potential h (0–1, in increments of 0.1) because the swan population used to sustain high rates of egg removal during the early and mid-20th century (Lamb 1964, Williams 1979). For each combination of SAHY and h, we held both of these rates constant and used n = 10,000 random samples from each of the bootstrap distributions of the other parameters described above to estimate λ. This approach allowed us to assess uncertainty in for each combination of SAHY and h.
RESULTS
The demographic data we used for modeling swan population dynamics were congruent with data from previous studies (Table 1; see Appendix 1), and the model provided ecologically plausible estimates of demographic parameters (Figs. 3, 4, Table 2).
Differential effects of demographic rates on population growth
The mean of the bootstrap distribution of λ was 1.018 (1.8% annual growth) (95% CI 0.808–1.241). Both sensitivity and elasticity analyses showed that λ was affected most strongly by changes in SAHY, followed by changes in SHY, b, and, least of all, f0 (Table 2, Fig. 3). Based on the means of the bootstrap distributions of sx and ex, the effect of an absolute change in SAHY on λ was about 30 times greater than that of f0, whereas the effect of a proportional change in SAHY on λ was almost six times greater than that of f0 (Table 2). Uncertainty in λ was almost entirely due to uncertainty in SHY and SAHY (Table 2).
Life expectancy and reproductive value of after-hatch-year swans
The model estimated that life expectancy of AHY swans was 5.7 years (95% CI: 3.4–17.0; see Appendix 1), and the mean number of fresh eggs and hatched eggs each AHY swan would produce in its lifetime, assuming no egg harvest pressure, were 5.9 (95% CI: 2.1–20.2) and 4.5 (95% CI: 1.7–15.3), respectively (Fig. 3).
Sustainability of mixed management
Assuming the mean of the bootstrap distribution of SAHY, the model estimated that the proportion of all swan nests at Te Waihora that could have all eggs from first clutches harvested sustainably was ~0.3 (Fig. 4). However, the level of egg harvest that the population could sustain depended strongly on SAHY (Fig. 4). For instance, the estimated proportion of all swan nests that could have first-clutch eggs harvested sustainably decreased to zero if SAHY decreased by only 0.02, and approximately doubled to ~0.6 if SAHY increased by 0.02 (Fig. 4). If SAHY increased by 0.04, the model estimated that harvesting almost all eggs from first clutches at Te Waihora would be sustainable (Fig. 4).
DISCUSSION
Sustainable species management requires understanding how populations respond to different demographic pressures and iteratively adapting management to population and environmental fluctuations (Moller et al. 2004, Weinbaum et al. 2013). The Te Waihora swan population was more sensitive to changes in survival of subadults and adults (AHY individuals) than to changes in fertility, consistent with other long-lived waterfowl species (Cooch et al. 2014, Koons et al. 2014, Wood et al. 2019). Moreover, population sensitivity to fertility was mostly due to underlying sensitivity to breeding propensity rather than nest hatching success. In addition, uncertainty in the current swan population growth rate was almost entirely attributable to uncertainty in swan survival rates after hatching (Table 2). These results could be particularly useful for informing potential efforts to enhance population monitoring and harvest management.
The limitations of our model reflect some of the challenges associated with collecting demographic data on animal populations. Specifically, indirect estimates of HY swan survival, and small sample size for estimates of AHY swan survival, resulted in large uncertainty in model estimates. More robust estimates of survival in long-lived species have been based on data from long-term monitoring (e.g., mark-recapture) programs that include hundreds or thousands of observations (Sandercock 2006). Such monitoring programs are costly to develop and maintain and have not been undertaken for swans in New Zealand in decades (Williams 1977, 1979, Barker and Buchanan 1993). Nevertheless, our estimates of swan survival were congruent with previous estimates for swans in New Zealand based on long-term mark-recapture datasets (Figs A1.3, Fig A1.5; Williams 1977, 1979, Barker and Buchanan 1993), and with estimates for other swan species globally (Bart et al. 1991), indicating that they are at least realistic. However, the lack of information on density dependence to incorporate into our model could have resulted in unrealistically high estimates of population growth under some modeled management scenarios (Fig. 4). For example, AHY survival and population growth are likely regulated at least partially by availability of macrophyte food, which is scarce at Te Waihora (Lineham 1983, Gerbeaux 1989, Ford et al. 2017) and could prevent AHY survival from increasing to some of the levels we considered, regardless of potential reductions in sport hunting. More robust data (e.g., from a long-term monitoring program) could improve the precision and reduce potential bias of our estimates and allow the inclusion of additional demographic parameters such as density dependence. In addition, data on finer-resolution age or stage classes could help to account for potential transient population dynamics (Koons et al. 2005, Ezard et al. 2010), which we found to be minor (Fig. A1.6). Despite these data limitations, our model was useful for assessing differential effects of age-specific demographic rates on population growth, and how offtake from sport hunting affects the sustainability of potential customary egg harvests by Ngāi Tahu.
Management implications
Our results raise immediate concerns about contemporary species management by FGNZ, which has implications for customary management by Ngāi Tahu. The Te Waihora swan population is highly sensitive to mortality of AHY swans, yet FGNZ manages swan numbers through permitted sport hunting of (mostly) AHY swans, regulated through fixed quotas. Even a slight over-estimation of the population’s capacity to sustain sport hunting, if not compensated for by increased survival and/or reproduction of remaining individuals, could lead to population declines (Pöysä et al. 2004, Sandercock et al. 2011) and preclude sustainable customary egg harvests. The risk of such an over-estimation is elevated because the swan hunting season (May–September) overlaps with the breeding season (peak nesting in August–September), and groups of hunters on farms surrounding Te Waihora sometimes shoot large numbers of swans (> 100) as they stage for nesting (Donald Brown, Ngāi Tahu, Christchurch, New Zealand, 9 February 2021, personal communication). This activity could have a disproportionately large impact on the population by reducing not only annual adult survival but also the proportion of surviving adults that attempt breeding, particularly if shooting during the pre-nesting period disrupts social cues that facilitate synchronous colonial breeding (Burger 1988, Reed and Dobson 1993, Wilhelm and Storey 2002). Tangata tiaki and customary egg harvesters have reported that the abundance of breeding swans at Te Waihora’s largest swan nesting colony has declined, which could be at least partly due to swans being killed immediately prior to, or during, nesting (Donald Brown and Craig Pauling, Ngāi Tahu, Christchurch, New Zealand, 9 February 2021, personal communication). Essentially, landowners are using the FGNZ gamebird regulations to control swan numbers in their area to limit potential swan impacts on new grass and crops. In addition, FGNZ does not monitor baseline mortality rates of swans or require landowners or gamebird license holders to report their offtake, as described above. Considering this study and related studies (Herse et al. 2020, 2021), and concerns expressed by local tangata tiaki, regional FGNZ managers recently indicated that they plan to adjust local gamebird regulations to reduce sport hunting pressure on swans at Te Waihora. However, lack of monitoring and resulting uncertainty in FGNZ management limit the utility of our demographic model, and the ability of tangata tiaki to assess sustainable rates of egg harvest within the current reporting environment. Thus, the need to monitor baseline rates of swan survival and effects of hunting more closely, to ensure that gamebird regulations are sustainable and do not preclude sustainable customary harvest, is a key recommendation to emerge from this study.
Fully recognizing the rights and interests of Ngāi Tahu, and empowering tangata tiaki authority in local species management, could lead to win-win outcomes that improve population and environmental monitoring (Herse et al. 2020) and, if/when appropriate, control swan numbers without wasting resources or killing swans unnecessarily (Bataille 2020, Bataille et al. 2021, Herse et al. 2021). Tangata tiaki and customary egg harvesters could enhance monitoring of swans and Te Waihora by continually generating information on breeding activity (e.g., locations and sizes of nesting colonies, hatching success) and environmental pressures (e.g., water pollution, flooding, nest depredation), which is currently lacking in local species management by FGNZ (Herse et al. 2020, 2021). Customary harvest could also be a practical, culturally appropriate, and less wasteful alternative to non-consumptive culling for controlling swan numbers to limit grazing of pasture in problem areas (Bataille 2020, Bataille et al. 2021, Herse et al. 2021). Tangata tiaki have rights to decide what level of swan numbers is appropriate to maintain (United Nations General Assembly 2007, Orange 2011), whereas current decision making is biased toward the interests of sport hunters and farmers (New Zealand Government 1987, Bataille 2020, Bataille et al. 2021). Although most landowners around Te Waihora value wetland health, active farmers generally prioritize economic gains, which motivates some to shoot swans to limit potential grazing impacts (Bataille 2020, Bataille et al. 2021). Ironically, grazing of pasture by swans can be partly attributed to macrophyte scarcity caused by pollution from farming (Lineham 1983, Gerbeaux 1989, Hearnshaw and Hughey 2010, Williams 2013a). Thus, efforts to reduce swan grazing impacts for farmers should prioritize curbing pollution that degrades Te Waihora and leads swans to feed on pasture (Williams 1979, 2013a), before resorting to local swan population control. However, if tangata whenua and the Crown make an equitable decision to control swan numbers, customary egg harvest could provide a fine-tune dial for managing population size, such that a potential over-harvest would pose less of a threat to the population when compared to an excess of sport hunting (or culling under the guise of gamebird regulations; Fig. 4). Harvesting from early life stages (eggs or juveniles) is a commonly used strategy by IPLC to fine-tune or reduce impacts of customary harvest (Gudmundsson 1979, Hunter et al. 2000, Lyver et al. 2008).
Caveats to the implementation and potential benefits of customary management
Supporting efforts by IPLC to engage with their environments, generate and maintain ecological knowledge, and practice customary management can improve local conservation and community livelihoods (Popp et al. 2019, Freitas et al. 2020). However, achieving the potential benefits of customary management requires addressing several related caveats. Here we focus on three caveats that are relevant to customary management by Ngāi Tahu and other IPLC elsewhere.
First, improving IPLC access to information about proposed or ongoing environmental management by non-IPLC groups could allow customary managers to make informed decisions, which are key to sustainability (Armitage 2005, Akins et al. 2019). For instance, in this study, the lack of available information on FGNZ-permitted sport hunting of swans creates uncertainty for tangata tiaki who wish to harvest eggs sustainably. Regional gamebird management (currently through FGNZ) could alleviate this uncertainty by monitoring baseline rates of swan survival and permitted mortality more closely, e.g., by establishing a band-recovery program (Williams 1979, Sandercock 2006), and sharing their findings with tangata tiaki. The model we developed could aid communication within a potential future co-management arrangement between tangata tiaki and representatives of New Zealand’s Crown government, within which participants could share information about the swan population and different ecological pressures. Such collaboration and sharing of knowledge among IPLC and non-IPLC groups can improve collective understanding of social-ecological systems, and sustainability in environmental management (Folke et al. 2005, Armitage et al. 2009, Popp et al. 2019, Wehi et al. 2019). Importantly, effective collaborations in environmental management require government support for adaptive policies that recognize IPLC rights to resources and decision making, monitoring efforts (e.g., band-recovery programs), and time to establish trust among different groups (Folke et al. 2005, Akins et al. 2019). Collaborations between IPLC and scientists (IPLC and non-IPLC) can also provide a pathway to partially fund the generation of new knowledge, e.g., using demographic models (Jones et al. 2015, McKechnie et al. 2020) and support customary management (Ens et al. 2016, Popp et al. 2019, Freitas et al. 2020).
Second, curbing human-driven environmental degradation is important to supporting customary management (Tang and Gavin 2016, Lyver et al. 2019b). For instance, agricultural pollution at Te Waihora over the past half-century has prevented growth of swans’ primary food source, macrophytes (Lineham 1983, Gerbeaux 1989), and apparently reduced swans’ propensity to lay replacement eggs following harvest (Williams 1979, Herse et al. 2021). Thus, regardless of pressure from sport hunting of swans, pollution at Te Waihora limits local Ngāi Tahu abilities to engage in sustainable egg harvests, through swans’ reduced resilience to egg harvest, and some egg harvesters’ fears of consuming eggs containing pollutants (Bataille 2020, Bataille et al. 2021). Over time, declines in tangata tiaki and customary harvester engagement with Te Waihora could lead to a loss of local knowledge about swans and their responses to egg harvest and other pressures. Because customary harvests and sharing of resources are central to the well-being of many IPLC (Tyrrell 2008, Caillon et al. 2017, Freitas et al. 2020), including Māori (Kawharu 2000, Bataille 2020, Bataille et al. 2021), reduced engagement in customary management can have cascading effects on cultural health and diversity (Lyver et al. 2019b, Fernández-Llamazares et al. 2021, Yletyinen et al. 2022). Thus, to alleviate these threats, we urge governments to adopt place-based environmental management policies that recognize links between environmental and cultural well-being (Gavin et al. 2015, Caillon et al. 2017, Akins et al. 2019, Lyver et al. 2019a).
Finally, improving social equity in environmental governance is imperative to achieving the potential benefits of customary management (Brondizio and Tourneau 2016, Ruru et al. 2017). Restrictive policies that neglect IPLC rights and values limit engagement and the continual generation and updating of knowledge (Stephenson et al. 2014, Parlee et al. 2018). Such policies can lead to inappropriate conservation approaches that are biased toward dominant cultures and against IPLC (Moller 1996, Tyrrell 2008, Eichler and Baumeister 2018, Parlee et al. 2018, Guibrunet et al. 2021). For example, New Zealand law fully protects most native bird species from offtake of any kind, regardless of their conservation status or value for Māori, yet classifies other species valued by sport hunters as gamebirds, including one with a conservation status of “nationally vulnerable” (pārera or Grey Duck, Anas superciliosa; New Zealand Government 1953, Williams 2013b, Wehi and Lord 2017, Lyver et al. 2019a). Even in the case of the Black Swan, a species commonly shot by sport hunters and farmers, tangata tiaki cannot authorize customary egg harvests without special permission from the Minister of Conservation or regional FGNZ managers, which is inconsistent with the Treaty, as described above, and creates a barrier for Ngāi Tahu engagement (Bataille 2020, Bataille et al. 2021). Putting aside the hypocrisy of restrictions on customary egg harvest (government-mandated gamebird managers destroyed and collected and sold swan eggs for decades during the 20th century while prohibiting customary harvest by Ngāi Tahu; Lamb 1964, Williams 1979, Pauling and Arnold 2008), the results of this study suggest that the long-term scrutiny of customary egg harvest is unfounded, considering the Te Waihora population is far more sensitive to AHY swan mortality than to changes in nest hatching success. Thus, implementing customary management requires confronting cultural biases and power asymmetries against IPLC, and adopting equitable policies that empower IPLC to engage with and protect their lands and resources in culturally appropriate ways (Pinkerton et al. 2014, Ruru et al. 2017, Wehi and Lord 2017).
In conclusion, we emphasize that engaging different cultures and perspectives in environmental management can improve knowledge production, adaptability, and sustainability within social-ecological systems, especially if individuals and groups share power, build trust, and find common grounds to work from, e.g., mutual interests in environmental stewardship (Folke et al. 2005, Armitage et al. 2009, Tengö et al. 2014, Bataille et al. 2021). However, differences in perspectives, priorities, abilities, expectations, and authority among individuals and groups can spur conflicts that sow division and mistrust and constrain management (Moller 1996, 1998, Coombes and Hill 2005, Brondizio et al. 2021). Averting such dilemmas is important to cooperation and sustainability and therefore requires steps to resolve and mediate conflicts (Coombes and Hill 2005, Brondizio et al. 2021), including recognition of the rights and aspirations of IPLC (United Nations General Assembly 2007, McGregor et al. 2020, Reyes-García et al. 2022), and establishment of collaborative cross-cultural learning and capacity-building initiatives (Whyte 2013, Tengö et al. 2014, Tipa et al. 2016). Reversing human-driven declines in biodiversity and ecosystem health will clearly not be easy, so we urge governments and environmental managers to pursue adaptive solutions that are both effective and socially equitable.
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.AUTHOR CONTRIBUTIONS
All authors contributed to the design of the study and the writing of the manuscript. M.R.H. led the data collection, modeling, and writing of the manuscript. N.J.S. and P.O.L. secured the primary funding for the study.
ACKNOWLEDGMENTS
We thank Ngāi Tahu Advisory Committee members Donald Brown, Iaean Cranwell, Cyril Gilroy, John Henry, David Higgins (chair), Jane Kitson, Craig Pauling, Makarini Rupene, Khyla Russell, Joe Taurima, and the late Paul Wilson for their expert advice and guidance. We thank pilots Ivor Yockney and Hugh Robinson, skipper Renny Bishop, and field assistants Sophie Allen, Alba Cervantes Loreto, Laís Maia, Michelle Marraffini, Oscar Pollard, Marcus Shadbolt, and Laura Young for their dedicated work. Chris Jones contributed to the initial design and development of the study. We thank Murray Williams for providing insights and information, and Lucas Pereira Martins and Brett Sandercock for providing R scripts that aided analyses. This project was funded by the Ministry of Business, Innovation and Employment’s New Zealand’s Biological Heritage National Science Challenge (Project 3.2), and Strategic Science Investment Fund.
DATA AVAILABILITY
The data and R code that support the findings of this study are jointly held by Te Rūnanga o Ngāi Tahu and Manaaki Whenua Landcare Research. The data are archived at MWLR (https://doi.org/10.34721/HPRD-HY85).
LITERATURE CITED
Adams, J. S. 1971. Black swans at Lake Ellesmere. Wildlife-a Review 3:23-25.
Akins, A., P. O’B. Lyver, H. F. Alrøe, and H. Moller. 2019. The universal precautionary principle: new pillars and pathways for environmental, sociocultural, and economic resilience. Sustainability 11(8):2357. https://doi.org/10.3390/su11082357
Armitage, D. 2005. Adaptive capacity and community-based natural resource management. Environmental Management 35(6):703-715. https://doi.org/10.1007/s00267-004-0076-z
Armitage, D. R., R. Plummer, F. Berkes, R. I. Arthur, A. T. Charles, I. J. Davidson-Hunt, A. P. Diduck, N. C. Doubleday, D. S. Johnson, M. Marschke, P. McConney, E. W. Pinkerton, and E. K. Wollenberg. 2009. Adaptive co-management for social-ecological complexity. Frontiers in Ecology and the Environment 7(2):95-102. https://doi.org/10.1890/070089
Artelle, K. A., M. Zurba, J. Bhattacharyya, D. E. Chan, K. Brown, J. Housty, and F. Moola. 2019. Supporting resurgent Indigenous-led governance: a nascent mechanism for just and effective conservation. Biological Conservation 240:108284. https://doi.org/10.1016/j.biocon.2019.108284
Ban, N. C., L. Eckert, M. McGreer, and A. Frid. 2017. Indigenous knowledge as data for modern fishery management: a case study of Dungeness crab in Pacific Canada. Ecosystem Health and Sustainability 3(8):1379887. https://doi.org/10.1080/20964129.2017.1379887
Barker, R. J., and I. M. Buchanan. 1993. Estimating adult survival rates of New Zealand Black Swans banded as immatures. Journal of Wildlife Management 57(3):549-555. https://doi.org/10.2307/3809281
Bart, J., S. Earnst, and P. J. Bacon. 1991. Comparative demography of the swans: a review. Pages 15-21 in J. Sears and P. J. Bacon, editors. Proceedings of the Third International Swan Symposium. Wildfowl Supplement No. 1. International Waterfowl and Wetlands Research, Slimbridge, UK.
Bataille, C. Y. 2020. Cross-cultural interest groups’ values and desired states inform biocultural management of wetlands in Aotearoa New Zealand. Dissertation. University of Canterbury, Christchurch, New Zealand.
Bataille, C. Y., S. K. Malinen, J. Yletyinen, N. Scott, and P. O’B. Lyver. 2021. Relational values provide common ground and expose multi-level constraints to cross-cultural wetland management. People and Nature 3(4):941-960. https://doi.org/10.1002/pan3.10244
Berkes, F. 2009. Indigenous ways of knowing and the study of environmental change. Journal of the Royal Society of New Zealand 39(4):151-156. https://doi.org/10.1080/03014220909510568
Bradshaw, C. J. A., and B. W. Brook. 2007. Ecological-economic models of sustainable harvest for an endangered but exotic megaherbivore in Northern Australia. Natural Resource Modeling 20(1):129-156. https://doi.org/10.1111/j.1939-7445.2007.tb00203.x
Brockington, D., and J. Igoe. 2006. Eviction for conservation: a global overview. Conservation & Society 4(3):424-470.
Brondizio, E. S., K. Andersson, F. de Castro, C. Futemma, C. Salk, M. Tengö, M. Londres, D. C. Tourne, T. S. Gonzalez, A. Molina-Garzón, G. Russo Lopes, and S. M. Siani. 2021. Making place-based sustainability initiatives visible in the Brazilian Amazon. Current Opinion in Environmental Sustainability 49:66-78. https://doi.org/10.1016/j.cosust.2021.03.007
Brondizio, E. S., and F.-M. L. Tourneau. 2016. Environmental governance for all. Science 352(6291):1272-1273. https://doi.org/10.1126/science.aaf5122
Brook, B. W., D. M. J. S. Bowman, C. J. A. Bradshaw, B. M. Campbell, and P. J. Whitehead. 2006. Managing an endangered Asian bovid in an Australian national park: the role and limitations of ecological-economic models in decision-making. Environmental Management 38(3):463-469. https://doi.org/10.1007/s00267-005-0157-7
Burger, J. 1988. Social attraction in nesting least tern: effects of numbers, spacing, and pair bonds. Condor 90(3):575-582. https://doi.org/10.2307/1368345
Caillon, S., G. Cullman, B. Verschuuren, and E. J. Sterling. 2017. Moving beyond the human-nature dichotomy through biocultural approaches: including ecological well-being in resilience indicators. Ecology and Society 22(4):27. https://doi.org/10.5751/ES-09746-220427
Caswell, H. 2001. Matrix population models: construction, analysis and interpretation. Second edition. Sinauer Associates, Sunderland, Massachusetts, USA.
Chaloupka, M., and G. Balazs. 2007. Using Bayesian state-space modelling to assess the recovery and harvest potential of the Hawaiian green sea turtle stock. Ecological Modelling 205(1-2):93-109. https://doi.org/10.1016/j.ecolmodel.2007.02.010
Cinner, J. E., and S. Aswani. 2007. Integrating customary management into marine conservation. Biological Conservation 140(3-4):201-216. https://doi.org/10.1016/j.biocon.2007.08.008
Cooch, E. G., M. Guillemain, G. S. Boomer, J.-D. Lebreton, and J. D. Nichols. 2014. The effects of harvest on waterfowl populations. Wildfowl Special Issue 4:220-227.
Coombes, B. L., and S. Hill. 2005. “Na whenua, na Tuhoe. Ko D.o.C. te partner”—Prospects for Comanagement of Te Urewera National Park. Society & Natural Resources 18(2):135-152. https://doi.org/10.1080/08941920590894516
Cumming, G. S., D. H. M. Cumming, and C. L. Redman. 2006. Scale mismatches in social-ecological systems: causes, consequences, and solutions. Ecology and Society 11(1):14. https://doi.org/10.5751/ES-01569-110114
Dawson, N. M., B. Coolsaet, E. J. Sterling, R. Loveridge, N. D. Gross-Camp, S. Wongbusarakum, K. K. Sangha, L. M. Scherl, H. Phuong Phan, N. Zafra-Calvo, W. G. Lavey, P. Byakagaba, C. J. Idrobo, A. Chenet, N. J. Bennett, S. Mansourian, and F. J. Rosado-May. 2021. The role of Indigenous peoples and local communities in effective and equitable conservation. Ecology and Society 26(3):19. https://doi.org/10.5751/ES-12625-260319
Díaz, S., J. Settele, E. S. Brondízio, H. T. Ngo, J. Agard, A. Arneth, P. Balvanera, K. A. Brauman, S. H. M. Butchart, K. M. A. Chan, L. A. Garibaldi, K. Ichii, J. Liu, S. M. Subramanian, G. F. Midgley, P. Miloslavich, Z. Molnár, D. Obura, A. Pfaff, S. Polasky, A. Purvis, J. Razzaque, B. Reyers, R. R. Chowdhury, Y.-J. Shin, I. Visseren-Hamakers, K. J. Willis, and C. N. Zayas. 2019. Pervasive human-driven decline of life on Earth points to the need for transformative change. Science 366(6471):eaax3100. https://doi.org/10.1126/science.aax3100
Eichler, L., and D. Baumeister. 2018. Hunting for justice: an Indigenous critique of the North American model of wildlife conservation. Environment and Society 9:75-90. https://doi.org/10.3167/ares.2018.090106
Ens, E., M. L. Scott, Yugul Mangi Rangers, C. Moritz, and R. Pirzl. 2016. Putting indigenous conservation policy into practice delivers biodiversity and cultural benefits. Biodiversity and Conservation 25(14):2889-2906. https://doi.org/10.1007/s10531-016-1207-6
Ens, E., G. M. Towler, C. Daniels, Yugul Mangi Rangers, Manqurrk Rangers. 2012. Looking back to move forward: collaborative ecological monitoring in remote Arnhem Land. Ecological Management & Restoration 13(1):26-35. https://doi.org/10.1111/j.1442-8903.2011.00627.x
Ezard, T. H. G., J. M. Bullock, H. J. Dalgleish, A. Millon, F. Pelletier, A. Ozgul, and D. N. Koons. 2010. Matrix models for a changeable world: the importance of transient dynamics in population management. Journal of Applied Ecology 47(3):515-523. https://doi.org/10.1111/j.1365-2664.2010.01801.x
Fernández-Llamazares, Á., D. Lepofsky, K. Lertzman, C. G. Armstrong, E. S. Brondizio, M. C. Gavin, P. O’B. Lyver, G. P. Nicholas, P. Pascua, N. J. Reo, V. Reyes-García, N. J. Turner, J. Yletyinen, E. N. Anderson, W. Balée, J. Cariño, D. M. David-Chavez, C. P. Dunn, S. C. Garnett, S. Greening (La'goot), S. Jackson (Niniwum Selapem), H. Kuhnlein, Z. Molnár, G. Odonne, G.-B. Retter, W. J. Ripple, L. Sáfián, A. S. Bahraman, M. Torrents-Ticó, and M. B. Vaughan. 2021. Scientists’ warning to humanity on threats to Indigenous and local knowledge systems. Journal of Ethnobiology 41(2):144-169. https://doi.org/10.2993/0278-0771-41.2.144
Folke, C., T. Hahn, P. Olsson, and J. Norberg. 2005. Adaptive governance of social-ecological systems. Annual Review of Environment and Resources 30:441-473. https://doi.org/10.1146/annurev.energy.30.050504.144511
Ford, D. E., F. D. Hughey, and K. Taylor. 2017. Te Waihora/Lake Ellesmere: state of the lake 2017. Report No. 3. Waihora Ellesmere Trust Technical, Christchurch, New Zealand.
Freitas, C. T., P. F. Macedo Lopes, J. V. Campos‐Silva, M. M. Noble, R. Dyball, and C. A. Peres. 2020. Co‐management of culturally important species: a tool to promote biodiversity conservation and human well‐being. People and Nature 2(1):61-81. https://doi.org/10.1002/pan3.10064
Gadgil, M., F. Berkes, and C. Folke. 1993. Indigenous knowledge for biodiversity conservation. Ambio 22(2-3):151-156.
Gavin, M. C., J. McCarter, A. Mead, F. Berkes, J. R. Stepp, D. Peterson, and R. Tang. 2015. Defining biocultural approaches to conservation. Trends in Ecology & Evolution 30(3):140-145. https://doi.org/10.1016/j.tree.2014.12.005
Gerbeaux, P. J. 1989. Aquatic plant decline in Lake Ellesmere: a case for macrophyte management in a shallow New Zealand lake. Dissertation. Lincoln College, Lincoln, New Zealand.
Goldman, M. J. 2011. Strangers in their own land: Maasai and wildlife conservation in Northern Tanzania. Conservation & Society 9(1):64-79.
Gudmundsson, F. 1979. The past status and exploitation of the Mývatn waterfowl populations. Oikos 32(1-2):232-249. https://doi.org/10.2307/3544230
Guerrero, A. M., R. R. J. McAllister, J. Corcoran, and K. A. Wilson. 2013. Scale mismatches, conservation planning, and the value of social-network analyses. Conservation Biology 27(1):35-44. https://doi.org/10.1111/j.1523-1739.2012.01964.x
Guibrunet, L., P . R. W. Gerritsen, J. A. Sierra‐Huelsz, A. C. Flores‐Díaz, E. García‐Frapolli, E. García‐-Serrano, U. Pascual, and P. Balvanera. 2021. Beyond participation: how to achieve the recognition of local communities’ value‐systems in conservation? Some insights from Mexico. People and Nature 3(3):528-541. https://doi.org/10.1002/pan3.10203
Halim, A. A., N. Othman, S. R. Ismail, J. A. Jawan, and N. N. Ibrahim. 2012. Indigenous knowledge and biodiversity conservation in Sabah, Malaysia. International Journal of Social Science and Humanity 2(2):159-163. https://doi.org/10.7763/IJSSH.2012.V2.88
Hearnshaw, E. J. S., and K. F. D. Hughey. 2010. A tolerance range approach for the investigation of values provided by Te Waihora/Lake Ellesmere. Land Environment and People Research Report No. 17. Lincoln University, Lincoln, New Zealand.
Herse, M. R., P. O’B. Lyver, N. J. Scott, A. R. McIntosh, S. C. Coats, A. M. Gormley, and J. M. Tylianakis. 2020. Engaging Indigenous peoples and local communities could alleviate scale mismatches in social-ecological systems. BioScience 70(8):699-707. https://doi.org/10.1093/biosci/biaa066
Herse, M. R., J. M. Tylianakis, N. J. Scott, D. Brown, I. Cranwell, J. Henry, C. Pauling, A. R. McIntosh, and P. O’B. Lyver. 2021. Effects of customary egg harvest regimes on hatching success of culturally important waterfowl species. People and Nature 3(2):499-512. https://doi.org/10.1002/pan3.10196
Hunn, E., D. Johnson, P. Russell, and T. Thornton. 2003. Huna Tlingit traditional environmental knowledge, conservation, and the management of a “wilderness” park. Current Anthropology 44(Supplement):79-103. https://doi.org/10.1086/377666
Hunter, C., H. Moller, and J. Kitson. 2000. Muttonbirder selectivity of Sooty Shearwater (titi) chicks harvested in New Zealand. New Zealand Journal of Zoology 27(4):395-414. https://doi.org/10.1080/03014223.2000.9518249
Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services (IPBES). 2019a. Summary for policymakers of the global assessment report on biodiversity and ecosystem services. S. Díaz, J. Settele, E. S. Brondízio, H. T. Ngo, M. Guèze, J. Agard, A. Arneth, P. Balvanera, K. A. Brauman, S. H. M. Butchart, K. M. A. Chan, L. A. Garibaldi, K. Ichii, J. Liu, S. M. Subramanian, G. F. Midgley, P. Miloslavich, Z. Molnár, D. Obura, A. Pfaff, S. Polasky, A. Purvis, J. Razzaque, B. Reyers, R. Roy Chowdhury, Y. J. Shin, I. J. Visseren-Hamakers, K. J. Willis, and C. N. Zayas, editors. IPBES secretariat, Bonn, Germany. https://doi.org/10.5281/zenodo.3553579
Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services (IPBES). 2019b. Indigenous peoples and local communities. IPBES secretariat, Bonn, Germany. https://ipbes.net/glossary/indigenous-peoples-local-communities
Jones, C. J., P. O’B. Lyver, J. Davis, B. Hughes, A. Anderson, and J. Hohapata-Oke. 2015. Reinstatement of customary seabird harvests after a 50-year moratorium. Journal of Wildlife Management 79(1):31-38. https://doi.org/10.1002/jwmg.815
Kawharu, I. H. 1989. Translation of the Treaty of Waitangi. Page 321 in I. H. Kawahu, editor. Maori and Pakeha perspectives of the Treaty of Waitangi. Oxford University Press, Auckland, New Zealand.
Kawharu, M. 2000. Kaitiakitanga: a Maori anthropological perspective of the Maori socio-environmental ethic of resource management. Journal of the Polynesian Society 109(4):349-370.
Kinan, I., and P. Dalzell. 2005. Sea turtles as a flagship species: different perspectives create conflicts in the Pacific Islands. Maritime Studies 3(2):195-212.
Koons, D. N., J. B. Grand, B. Zinner, and R. F. Rockwell. 2005. Transient population dynamics: relations to life history and initial population state. Ecological Modelling 185(2-4):283-297. https://doi.org/10.1016/j.ecolmodel.2004.12.011
Koons, D. N., G. Gunnarsson, J. A. Schmutz, and J. J. Rotella. 2014. Drivers of waterfowl population dynamics: from teal to swans. Wildfowl Special Issue 4:169-191.
Lamb, R. C. 1964. Birds, beasts & fishes: the first hundred years of the North Canterbury Acclimatisation Society. North Canterbury Acclimatisation Society, Christchurch, New Zealand.
Leslie, P. H. 1945. On the use of matrices in certain population mathematics. Biometrika 33(3):183-212. https://doi.org/10.1093/biomet/33.3.183
Lineham, I. 1983. Eutrophication of Lake Ellesmere: a study of phytoplankton. Dissertation. University of Canterbury, Christchurch, New Zealand.
Lyver, P. O’B., J. Davis, L. Ngamane, L. Anderson, and P. Clarkin. 2008. Hauraki Maori Matauranga for the conservation and harvest of Titi, Pterodroma macroptera gouldi. Papers and Proceedings of the Royal Society of Tasmania 142(1):149-159. https://doi.org/10.26749/rstpp.142.1.149
Lyver, P. O’B., J. Ruru, N. J. Scott, J. M. Tylianakis, J. Arnold, S. K. Malinen, C. Y. Bataille, M. R. Herse, C. J. Jones, A. M. Gormley, D. A. Peltzer, Y. Taura, P. Timoti, C. Stone, M. Wilcox, and H. Moller. 2019a. Building biocultural approaches into Aotearoa - New Zealand’s conservation future. Journal of the Royal Society of New Zealand 49(3):394-411. https://doi.org/10.1080/03036758.2018.1539405
Lyver, P. O’B., P. Timoti, T. Davis, and J. M. Tylianakis. 2019b. Biocultural hysteresis inhibits adaptation to environmental change. Trends in Ecology & Evolution 34(9):771-780. https://doi.org/10.1016/j.tree.2019.04.002
Lyver, P. O’B., P. Timoti, S. J. Richardson, and A. M. Gormley. 2021. Alignment of ordinal and quantitative species abundance and size indices for the detection of shifting baseline syndrome. Ecological Applications 31(4):e02301. https://doi.org/10.1002/eap.2301
Lyver, P. O’B., and J. M. Tylianakis. 2017. Indigenous peoples: conservation paradox. Science 357(6347):142-143. https://doi.org/10.1126/science.aao0780
McGregor, D., S. Whitaker, and M. Sritharan. 2020. Indigenous environmental justice and sustainability. Current Opinion in Environmental Sustainability 43:35-40. https://doi.org/10.1016/j.cosust.2020.01.007
McKechnie, S., D. Fletcher, J. Newman, C. Bragg, P. W. Dillingham, R. Clucas, D. Scott, S. Uhlmann, P. O’B. Lyver, A. M. Gormley, Rakiura Tītī Islands Administering Body, and H. Moller. 2020. Separating the effects of climate, bycatch, predation and harvesting on tītī (Ardenna grisea) population dynamics in New Zealand: a model-based assessment. PLoS ONE 15(12):e0243794. https://doi.org/10.1371/journal.pone.0243794
Meis, T. 1988. Waterfowl management in North Canterbury. Thesis. University of Canterbury, Christchurch, New Zealand.
Moller, H. 1996. Customary use of indigenous wildlife - towards a bicultural approach to conserving New Zealand’s biodiversity. Pages 89-125 in B. McFagen and P. Simpson, editors. Biodiversity: papers from a seminar series on biodiversity. Science and Research Division, Department of Conservation, Wellington, New Zealand.
Moller, H. 1998. From mutual mistrust to co-management. Fish and Game New Zealand Newsletter 6:17-24.
Moller, H., F. Berkes, P. O’B. Lyver, and M. Kislalioglu. 2004. Combining science and traditional ecological knowledge: monitoring populations for co-management. Ecology and Society 9(3):2. https://doi.org/10.5751/ES-00675-090302
Montano, V., W. F. D. van Dongen, M. A. Weston, R. A. Mulder, R. W. Robinson, M. Cowling, and P.-J. Guay. 2018a. A genetic assessment of the human-facilitated colonization history of Black Swans in Australia and New Zealand. Evolutionary Applications 11(3):364-375. https://doi.org/10.1111/eva.12535
Montano, V., W. F. D. van Dongen, M. A. Weston, R. A. Mulder, R. W. Robinson, M. Cowling, and P.-J. Guay. 2018b. Response to Rawlence et al. (2017): Native or not? Extinct and extant DNA of New Zealand Black Swans. Evolutionary Applications 11(3):378-379. https://doi.org/10.1111/eva.12587
New Zealand Government. 1953. Wildlife Act 1953. Public Act 1953 No 31. New Zealand Government, Wellington, New Zealand.
New Zealand Government. 1987. Conservation Act 1987. Public Act 1987 No 65. New Zealand Government, Wellington, New Zealand. https://www.legislation.govt.nz/act/public/1987/0065/latest/DLM103610.html
New Zealand Government. 1998. Ngāi Tahu Claims Settlement Act 1998. Public Act 1998 No 97. New Zealand Government, Wellington, New Zealand. https://www.legislation.govt.nz/act/public/1998/0097/latest/whole.html
Orange, C. 2011. The Treaty of Waitangi. Second edition. Bridget Williams Books, Wellington, New Zealand. https://doi.org/10.7810/9781877242489
Parlee, B. L., J. Sandlos, and D. C. Natcher. 2018. Undermining subsistence: barren-ground caribou in a “tragedy of open access.” Science Advances 4(2):e1701611. https://doi.org/10.1126/sciadv.1701611
Pauling, C., and J. Arnold. 2008. Cultural health of the lake. Pages 77-83 in K. Hughey and K. Taylor, editors. Te Waihora/Lake Ellesmere: state of the lake and future management. Lincoln University, Lincoln, New Zealand.
Peery, M. Z., B. H. Becker, and S. R. Beissinger. 2006. Combining demographic and count-based approaches to identify source-sink dynamics of a threatened seabird. Ecological Applications 16(4):1516-1528. https://doi.org/10.1890/1051-0761(2006)016[1516:CDACAT]2.0.CO;2
Pinkerton, E., E. Angel, N. Ladell, P. Williams, M. Nicolson, J. Thorkelson, and H. Clifton. 2014. Local and regional strategies for rebuilding fisheries management institutions in coastal British Columbia: what components of comanagement are most critical? Ecology and Society 19(2):72. https://doi.org/10.5751/ES-06489-190272
Popp, J. N., P. Priadka, and C. Kozmik. 2019. The rise of moose co-management and integration of Indigenous knowledge. Human Dimensions of Wildlife 24(2):159-167. https://doi.org/10.1080/10871209.2019.1545953
Pöysä, H., J. Elmberg, P. Nummi, and K. Sjöberg. 2004. Ecological basis of sustainable harvesting: Is the prevailing paradigm of compensatory mortality still valid? Oikos 104(3):612-615. https://doi.org/10.1111/j.0030-1299.2004.12788.x
R Core Team. 2021. R: A language environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria.
Rawlence, N. J., A. Kardamaki, L. J. Easton, A. J. D. Tennyson, R. P. Scofield, and J. M. Waters. 2017. Ancient DNA and morphometric analysis reveal extinction and replacement of New Zealand’s unique Black Swans. Proceedings of the Royal Society B: Biological Sciences 284(1859):20170876. https://doi.org/10.1098/rspb.2017.0876
Rawlence, N. J., A. Kardamaki, L. J. Easton, A. J. D. Tennyson, R. P. Scofield, and J. M. Waters. 2018. Native or not? Ancient DNA rejects persistence of New Zealand’s endemic Black Swan: a reply to Montano et al. Evolutionary Applications 11(3):376-377. https://doi.org/10.1111/eva.12577
Rayne, A., G. Byrnes, L. Collier-Robinson, J. Hollows, A. McIntosh, M. Ramsden, M. Rupene, P. Tamati-Elliffe, C. Thoms, and T. E. Steeves. 2020. Centring Indigenous knowledge systems to re-imagine conservation translocations. People and Nature 2(3):512-526. https://doi.org/10.1002/pan3.10126
Reed, J. M., and A. P. Dobson. 1993. Behavioural constraints and conservation biology: conspecific attraction and recruitment. Trends in Ecology & Evolution 8(7):253-256. https://doi.org/10.1016/0169-5347(93)90201-Y
Reyes-García, V., Á. Fernández-Llamazares, Y. Aumeeruddy-Thomas, P. Benyei, R. W. Bussmann, S. K. Diamond, D. García-del-Amo, S. Guadilla-Sáez, N. Hanazaki, N. Kosoy, M. Lavides, A. C. Luz, P. McElwee, V. J. Meretsky, T. Newberry, Z. Molnár, I. Ruiz-Mallén, M. Salpeteur, F. S. Wyndham, F. Zorondo-Rodriguez, and E. S. Brondizio. 2022. Recognizing Indigenous peoples’ and local communities’ rights and agency in the post-2020 Biodiversity Agenda. Ambio 51:84-92. https://doi.org/10.1007/s13280-021-01561-7
Ruru, J., P. O’B. Lyver, N. J. Scott, and D. Edmunds. 2017. Reversing the decline in New Zealand’s biodiversity: empowering Māori within reformed conservation law. Policy Quarterly 13(2):65-71. https://doi.org/10.26686/pq.v13i2.4657
Sandercock, B. K. 2006. Estimation of demographic parameters from live-encounter data: a summary review. Journal of Wildlife Management 70(6):1504-1520. https://doi.org/10.2193/0022-541X(2006)70[1504:EODPFL]2.0.CO;2
Sandercock, B. K., E. B. Nilsen, H. Brøseth, and H. C. Pedersen. 2011. Is hunting mortality additive or compensatory to natural mortality? Effects of experimental harvest on the survival and cause-specific mortality of Willow Ptarmigan. Journal of Animal Ecology 80(1):244-258. https://doi.org/10.1111/j.1365-2656.2010.01769.x
Sheil, D., M. Boissière, and G. Beaudoin. 2015. Unseen sentinels: local monitoring and control in conservation’s blind spots. Ecology and Society 20(2):39. https://doi.org/10.5751/ES-07625-200239
Snook, J., A. Cunsolo, D. Borish, C. Furgal, J. D. Ford, I. Shiwak, C. T. R. Flowers, and S. L. Harper. 2020. “We’re made criminals just to eat off the land”: colonial wildlife management and repercussions on Inuit well-being. Sustainability 12(19):8177. https://doi.org/10.3390/su12198177
Sobrevila, C. 2008. The role of Indigenous peoples in biodiversity conservation: the natural but often forgotten partners. World Bank Group Report No. 44300. World Bank Group, Washington, D.C., USA.
Solomon, M. 2014. Locked out of national parks: a call to action from Kaiwhakahaere Tā Mark Solomon. Te Karaka 63:14-17. https://ngaitahu.iwi.nz/our_stories/locked-national-parks/
Stephenson, J., F. Berkes, N. J. Turner, and J. Dick. 2014. Biocultural conservation of marine ecosystems: examples from New Zealand and Canada. Indian Journal of Traditional Knowledge 13(2):257-265.
Stubben, C., B. Milligan, and P. Nantel. 2020. popbio: Construction and analysis of matrix population models, version 2.7, https://cran.r-project.org/web/packages/popbio/index.html
Tang, R., and M. C. Gavin. 2016. A classification of threats to traditional ecological knowledge and conservation responses. Conservation and Society 14(1):57-70.
Te Rūnanga o Ngāi Tahu, and Department of Conservation. 2005. Te Waihora Joint Management Plan: Mahere Tukutahi o Te Waihora. Te Rūnanga o Ngāi Tahu, and Department of Conservation, Christchurch, New Zealand. https://api.ecan.govt.nz/TrimPublicAPI/documents/download/2738434
Tengö, M., E. S. Brondizio, T. Elmqvist, P. Malmer, and M. Spierenburg. 2014. Connecting diverse knowledge systems for enhanced ecosystem governance: the multiple evidence base approach. Ambio 43(5):579-591. https://doi.org/10.1007/s13280-014-0501-3
Tipa, G., G. R. Harmsworth, E. Williams, and J. C. Kitson. 2016. Integrating mātauranga Māori into freshwater management, planning and decision-making. Pages 613-637 in P. G. Jellyman, T. Davie, C. P. Pearson, and J. S. Harding, editors. Advances in New Zealand freshwater science. New Zealand Hydrological Society, Wellington, New Zealand.
Tyrrell, M. 2008. Nunavik Inuit perspectives on beluga whale management in the Canadian Arctic. Human Organization 67(3):322-334. https://doi.org/10.17730/humo.67.3.47826252k0623352
United Nations General Assembly. 2007. United Nations Declaration on the Rights of Indigenous Peoples (No. A/RES/61/295). United Nations General Assembly, New York, New York, USA.
Wehi, P. M., J. R. Beggs, and T. G. McAllister. 2019. Ka mua, ka muri: the inclusion of mātauranga Māori in New Zealand ecology. New Zealand Journal of Ecology 43(3):3379.
Wehi, P. M., and J. M. Lord. 2017. Importance of including cultural practices in ecological restoration. Conservation Biology 31(5):1109-1118. https://doi.org/10.1111/cobi.12915
Weinbaum, K. Z., J. S. Brashares, C. D. Golden, and W. M. Getz. 2013. Searching for sustainability: are assessments of wildlife harvests behind the times? Ecology Letters 16(1):99-111. https://doi.org/10.1111/ele.12008
Whyte, K. P. 2013. On the role of traditional ecological knowledge as a collaborative concept: a philosophical study. Ecological Processes 2:7. https://doi.org/10.1186/2192-1709-2-7
Wilhelm, S. I., and A. E. Storey. 2002. Influence of cyclic pre-lay attendance on synchronous breeding in Common Murres. Waterbirds 25(2):156-163. https://doi.org/10.1675/1524-4695(2002)025[0156:IOCPAO]2.0.CO;2
Williams, M. 1977. Locations of recoveries of Black Swans, Cygnus atvatus Latham, Banded at Lake Whangape and Lake Ellesmere, New Zealand. Australian Wildlife Research 4:289-299. https://doi.org/10.1071/WR9770289
Williams, M. 1979. Status and management of Black Swans Cygnus atratus, Latham at Lake Ellesmere since the “Wahine” Storm, April 1968. New Zealand Journal of Ecology 2:34-41.
Williams, M. 1981. The demography of New Zealand’s Cygnus atratus population. Pages 147-160 in G. V. T. Matthews and M. Smart, editors. Proceedings of the Second International Swan Symposium. International Waterfowl and Wetlands Bureau, Slimbridge, UK.
Williams, M. 2013a. Black Swan | Kakīānau. In C. M. Miskelly, editor. New Zealand Birds Online. https://nzbirdsonline.org.nz/species/black-swan
Williams, M. 2013b [updated 2022]. Grey Duck | Pārera. In C. M. Miskelly, editor. New Zealand Birds Online. https://nzbirdsonline.org.nz/species/grey-duck
Witter, R., K. R. M. Suiseeya, R. L. Gruby, S. Hitchner, E. M. Maclin, M. Bourque, and J. P. Brosius. 2015. Moments of influence in global environmental governance. Environmental Politics 24(6):894-912. https://dx.doi.org/10.1080/09644016.2015.1060036
Wood, K. A., L. Cao, P. Clausen, C. R. Ely, L. Luigujõe, E. C. Rees, J. Snyder, D. V. Solovyeva, and R. Włodarczyk. 2019. Current trends and future directions in swan research: insights from the 6th International Swan Symposium. Wildfowl Special Issue 5:1-34.
Yletyinen, J., J. M. Tylianakis, C. Stone, and P. O’B. Lyver. 2022. Potential for cascading impacts of environmental change and policy on indigenous culture. Ambio 51:1110-1122. https://doi.org/10.1007/s13280-021-01670-3
Zador, S. G., J. F. Piatt, and A. E. Punt. 2006. Balancing predation and egg harvest in a colonial seabird: a simulation model. Ecological Modelling 195(3-4):318-326. https://doi.org/10.1016/j.ecolmodel.2005.11.002
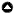
Fig. 1

Fig. 1. Map of our study area at Te Waihora/Lake Ellesmere, home to the largest population of Black Swans (Cygnus atratus) on the South Island of Aotearoa/New Zealand. Purple boxes outline swan breeding areas where we used orthophotography to census nests. Black and yellow circles indicate the current locations of the largest two swan breeding colonies at Te Waihora, where the data on nest fertility and egg harvest that we used in models originate. The map is a modified version of the NZ Topographic Basemap available on ArcGIS 10.5 (Environmental Systems Research Institute, Redlands, CA, USA; ©Eagle Technology Group Ltd.).
Fig. 2

Fig. 2. Life-cycle diagram of the age-classified matrix model we used to describe Black Swan (Cygnus atratus) population dynamics at Te Waihora/Lake Ellesmere, Aotearoa/New Zealand. Circles represent age classes (HY = hatch year, age < 1 year since hatching; AHY = after hatch year, age ≥ 1 year since hatching), whereas arrows represent annual per-capita demographic transition rates (FAHY = annual fertility or number of eggs hatched per AHY swan; SHY = probability of surviving the first year after hatching; SAHY = annual survival probability for AHY swans).
Fig. 3

Fig. 3. Flow diagram illustrating the steps we followed to develop the matrix model and estimate demographic parameters for the Black Swan (Cygnus atratus) population at Te Waihora/Lake Ellesmere, Aotearoa/New Zealand. First, we used empirical data to define initial bootstrap sampling distributions for population growth (λ), underlying parameters of annual fertility per after-hatch-year (AHY) swan (FAHY), and annual survival probability for AHY swans (SAHY). Second, we used random sampling from initial distributions to back-calculate, and hence generate a bootstrap distribution for, survival probability of swans over their first (hatch) year (SHY). Back-calculations of SHY incorporated information on survival of swan hatchlings (Shatchling), and adjusted the distributions for SAHY and underlying parameters of FAHY. Last, we used random sampling from the resulting bootstrap distributions to parameterize the matrix model and generate bootstrap distributions for a suite of demographic parameters for the focal population.
Fig. 4

Fig. 4. Heatmaps showing how changes in annual survival probability for after-hatch-year Black Swans (SAHY, Cygnus atratus), and nest harvest pressure (h), influence population growth rate (λ) at Te Waihora/Lake Ellesmere, Aotearoa/New Zealand. Warm colors indicate a declining population (λ < 1), cool colors indicate a growing population (λ > 1), and neutral white indicates a stable population (λ = 1). The middle plot shows the mean of a bootstrap distribution of λ for each combination of SAHY and h, whereas the left and right plots show the 25th and 75th percentile, respectively, for each combination.
Table 1
Table 1. Means and associated 95% confidence limits (in parentheses) of bootstrap distributions of demographic rates used to model Black Swan (Cygnus atratus) population dynamics at Te Waihora/Lake Ellesmere, Aotearoa/New Zealand. Demographic rates include population growth rate (λ), fertility (number of eggs hatched) per unharvested (f0) and harvested (fa) nest, total number of nests (Nnest), total number of after-hatch-year or AHY swans (NAHY), survival probability of swan hatchlings over their first 40 days of life (Shatchling), survival probability over the first (hatch) year (SHY), and annual survival probabiliy for AHY swans (SAHY). Distributions marked with subscript “a” were used to back-calculate SHY, and adjusted based on constraints related to Shatchling (see main text). Distributions marked with subscript “b” were used to parameterize the matrix model, whereas distributions marked with an asterisk were adjusted but not used in the matrix model.
| Demographic rate | Inititial bootstrap distribution | New or adjusted bootstrap distribution |
| λ | 1.020 (0.890, 1.170)a | 1.017 (0.898, 1.150)* |
| f0 | 4.64 (3.55, 5.83)a | 4.68 (3.60, 5.88)b |
| fa | 2.67 (1.64, 3.83)b | - |
| Nnest | 856 (800, 913)a | 856 (799, 912)b |
| NAHY | 5669 (5521, 5815)a | 5670 (5520, 5815)b |
| Shatchling | 0.68 (0.54, 0.80)a | 0.68 (0.54, 0.80)* |
| SHY | - | 0.33 (0.06, 0.66)b |
| SAHY | 0.82 (0.65, 1.0)a | 0.84 (0.71, 0.94)b |
Table 2
Table 2. Means and associated 95% confidence limits (in parentheses) of bootstrap distributions for different parameters corresponding with demographic rates (f0 is fertility or number of eggs hatched per unharvested nest; b is annual breeding propensity; SHY is the probability of surviving the first or hatch year; SAHY is annual survival probability for AHY swans) that determined estimated Black Swan (Cygnus atratus) population growth (λ) at Te Waihora/Lake Ellesmere, Aotearoa/New Zealand. ‘Sensitivity’ and ‘elasticity’ are measures of the effects of absolute and proportional changes, respectively, in a given demographic rate on λ. To assess sources of uncertainy in λ, we quantified the ‘contribution’ of variance in each demographic rate to total variance in λ. ‘Reproductive value’ refers to estimates of the expected total number of fresh eggs and hatched eggs (i.e., HY swans) produced by an individual AHY swan during its lifetime, on average. All estimates are based on a model that assumes no egg harvest pressure (h = 0); this special case of the model leads to the elasticities for f0, b, and SHY being identical.
| Demographic rate | Sensitivity of λ | Elasticity of λ | Contribution to uncertainty in λ |
| f0 | 0.032 (0.008, 0.056) | 0.145 (0.041, 0.229) | 0.027 |
| b | 0.502 (0.120, 0.886) | 0.145 (0.041, 0.229) | 0.001 |
| SHY | 0.499 (0.361, 0.694) | 0.145 (0.041, 0.229) | 0.530 |
| SAHY | 1.037 (1.002, 1.097) | 0.855 (0.771, 0.959) | 0.442 |