






The following is the established format for referencing this article:
Slauson, K., B. Howard, A. M. White, C. Maxwell, and T. Holland. 2022. Evaluating the effects of alternative landscape management scenarios on three old-forest-associated predators over 100 years in the fire-prone forests of the Sierra Nevada, USA. Ecology and Society 27(3):28.ABSTRACT
Developing and implementing landscape-scale management strategies capable of balancing the need for restoring natural fire regimes and promoting ecosystem resiliency for future climate change remains an urgent need globally. To help guide development of a landscape management strategy capable of meeting multiple objectives, five alternative landscape management scenarios for reducing the risk of uncharacteristically severe wildfire using thinning or managed and prescribed wildfire were developed by local forest managers in the Lake Tahoe region of the Sierra Nevada mountains. Effects of each scenario on forest structure, composition, and wildfire behavior were simulated over a 100-year period using the dynamic landscape simulation model LANDIS-II. We developed empirical territory occurrence models for three old-forest-associated predators, using 22 California Spotted Owl, 28 Northern Goshawk, and 16 female Pacific marten territories and presence-only modeling to evaluate the effects of each management scenario. The recruitment of more old-forest habitat across the simulated landscape was a more significant factor than any differences in the management scenarios, resulting in increases in the numbers of territories for all three predators, regardless of scenario. Increases in the numbers of territories were slowed in Scenario 4, which had the greatest amount of thinning, but the positive trend continued decades beyond when the other scenarios began to show a decline in territory numbers from severe wildfire. However, increases in the numbers of territories over time were slowed and overall were the lowest for the two old-forest predators that were most sensitive to the amount of old forest at the territory scale in the scenario with the greatest pace and scale of treatments. This suggests a trade-off between slowing the increase in the numbers of territories in the short term from forest growth by using fuels treatments with increased pace and scale to create forest structure that is less susceptible to severe wildfire in the long term, and that these management scenarios may need to be re-evaluated in 50 years before committing to continuing management efforts that may ultimately become detrimental for old-forest predators within 100 years.INTRODUCTION
The need to develop and implement landscape-scale management strategies capable of balancing the need to restore natural fire regimes and promote ecosystem resiliency for future climate change remains a major challenge globally. Forest managers in North America are challenged by the need to balance potentially competing objectives of reducing the risk of uncharacteristically severe wildfire and maintaining viable populations of species of conservation concern. Most forests in western North America evolved with fire regimes characterized by frequent, low- to moderate-severity wildfire that removed fuels such as shrubs, downed wood, and small trees (Agee 1993). However, decades of fire suppression have allowed unnaturally high accumulations of these surface and ladder fuels to form, which has increased the frequency and extent of high-severity wildfire that now threatens ecological and human communities (Miller et al. 2009). In addition, warmer and drier conditions associated with climate change may lead to increases in fire activity over the next century (Westerling and Bryant 2008). In response, landscape-scale fuels reduction treatments have been proposed to remove surface and ladder fuels, with the goal of reducing uncharacteristically severe wildfire (USFS 2004).
Fuels treatments have the potential to negatively impact wildlife species associated with old-forest conditions because they change forest structure in ways that may affect survival and reproduction. For example, reproduction by the California Spotted Owl (Strix occidentalis occidentalis) has been shown to be negatively impacted by timber harvests that are characteristic of mechanical fuels treatments (Tempel et al. 2014), and landscape-scale mechanical fuels treatments have caused declines in territory occupancy (Stephens et al. 2014). Mechanical fuels treatments that simplify stand structure have also negatively affected movements of and habitat connectivity for the Pacific marten (Martes caurina) (Moriarty et al. 2016). The Northern Goshawk (Accipiter gentilis) is considered to be sensitive to changes in forest structure and composition from timber harvests in western forests (Reynolds et al. 1992), and has been shown to have lower territory occupancy and reproduction in response to timber harvesting (Crocker-Bedford 1990). While fuels treatments have the potential to benefit old-forest-associated species over the long term by reducing the risk of habitat loss from uncharacteristically severe wildfire (Collins et al. 2011, Scheller et al. 2011), forest managers must carefully design landscape management strategies with treatment prescriptions and spatial distributions that are implemented at rates capable of balancing the risks of short-term impacts with longer term benefits.
In 2017, the Lake Tahoe West Restoration Partnership began with the goal of developing and evaluating alternative landscape management approaches for the Lake Tahoe Basin. This effort included land managers, individuals interested in management of the Lake Tahoe Basin, and a multidisciplinary science team. The Lake Tahoe Basin is composed primarily of upper elevation montane and subalpine forest that is managed by the U.S. Forest Service but has a mosaic of private and state lands intermixed. The role of land managers was to develop a set of plausible management scenarios to be simulated over a 100-year period to understand their effects on a diverse set of indicators, including (1) risk of severe wildfire, (2) biodiversity and old-forest-associated predators, (3) air quality, (4) lake clarity, and (5) economics. The science team’s role was to measure the effects of the outcomes from each management scenario on each indicator. Changes in landscape vegetation structure and composition over the 100-year simulation period were modeled using LANDIS-II (https://www.landis-ii.org/), a spatial landscape simulation model designed to simulate forest succession and disturbances (Scheller et al. 2007).
Our overall objective was to develop empirical models for three old-forest-associated predators that are broadly of concern throughout forested landscapes in the west. We estimated changes in the number and distribution of occupied territories for each species in response to changes in landscape composition in five alternative management scenarios. The California Spotted Owl (owl), Pacific marten (marten), and Northern Goshawk (goshawk) are associated with old-forest conditions, and are species of conservation and management concern in the Lake Tahoe Basin. To develop territory occurrence models, we used known locations of owl and goshawk territories that were detected over three decades of surveys conducted throughout the Lake Tahoe Basin. Unlike the social structure of the owl and goshawk, where males and females form pairs and share efforts in raising their young, the marten has a polygamous mating system in which females are solely responsible for raising young and defend territories only against same-sex conspecifics (Powell 1994). Therefore, reproduction is tied to female territories, and prior research in the Lake Tahoe Basin has shown that males are more numerous, wide ranging, and use a wider variety of habitat conditions than females (Slauson 2017). Therefore, we used only known female territories to develop the marten territory model. Each species’ model was developed using a combination of candidate biophysical variables that would not change and variables related to forest structure and composition that would change during the simulation period. Our final objective was to evaluate the effect of each management scenario on each old-forest-associated predator by using the models to estimate changes in the number of occupied territories over the 100-year simulation period.
METHODS
Study area
This study was conducted in the ~80,000-ha Lake Tahoe Basin, nestled between the central Sierra Nevada mountains of California and the Carson Range of Nevada, USA. Elevations ranged from approximately 2000 to 3000 m, and the area is composed largely of forested habitats dominated by red fir (Abies magnifica), lodgepole pine (Pinus contorta), white fir (A. concolor), western white pine (P. monticola), mountain hemlock (Tsuga mertensiana), and Jeffrey pine (P. jeffreyi). The Lake Tahoe Basin has a dry-summer continental climate with average temperatures ranging between 25.9° and 4.3°C, and an average of 1440 mm of precipitation, which falls predominantly as snow.
Modeling changes in landscape composition
Baseline vegetation conditions for 2010 were created using a raster with 1-ha pixels across the non-lake portion of the Lake Tahoe Basin and input into the LANDIS-II spatial landscape change model following the methods of Loudermilk et al. (2013). The model then uses individual sub-models for species-specific growth to represent succession (Scheller et al. 2007), forest harvest and fuels treatments (Syphard et al. 2011), insect mortality (Sturtevant et al. 2004), and wildfire from both natural and anthropogenic ignitions. The model explicitly incorporates variation in fire behavior and effects on vegetation (Scheller et al. 2019) to simulate changes in landscape structure and composition annually over a 100-year period. To apply the spatially explicit fuels treatment alternatives for each management scenario, the Lake Tahoe Basin was divided into four management zones: (1) wildland-urban interface defense (WUI-defense [35%]), (2) wildland-urban interface threat (WUI-threat [29%]), (3) general forest (27%), and (4) designated wilderness areas (9%).
Management Scenario 1 included 100% fire suppression only. Scenario 2 was a WUI-focused alternative consistent with ongoing fuels reduction targets of 75% (300 ha/year) in the WUI-defense and 25% (100 ha/year) in the WUI-threat zones. Scenario 3 increased the pace (1600 ha/year) and scale (WUI, general forest, and wilderness) of fuels treatments, with 45% in the WUI-defense zone (725 ha/year), 25% in both the WUI-threat and general forest zones (400 ha in each zone/year), and 5% in the wilderness zone (80 ha/year). Scenario 4 further increased the pace (1600 ha/year) and scale (WUI, general forest, and wilderness) of fuels treatments with the addition of managed wildfire and prescribed fire (223 ha/year); thinning treatments occurred only in the WUI and were the same as those in Scenario 2. Suppression of wildfire remained at 100% in the WUI-defense zone, but managed wildfire was allowed in all other zones, and prescribed fire was allowed in all zones, with a goal of burning 1300 ha at low to moderate severity annually. Scenario 5 was a fire-focused strategy that combined the modest WUI thinning of Scenario 4 with much greater use of prescribed burning in all zones (425 ha/year) and managed natural ignitions for resource objectives in the general forest and wilderness (see Maxwell et al. 2022 for more details on management prescriptions and disturbance processes). Each year of simulation was replicated 10 times to provide an annual range of variability, and used a moderate projection of climate change conditions. To evaluate the effects of management scenarios on old-forest-associated predators, we used the 10 replicates for each 20-year time step, with year 0 representing baseline conditions and years 20, 40, 60, 80, and 100 serving as the temporal points for measuring changes.
Overall old-forest predator modeling approach
Due to the management and conservation status of California Spotted Owl and Northern Goshawk, data sets of the distribution of known territories detected during standardized survey protocols conducted from 1987 to 2017 were available. However, while these data sets included the spatial locations of nests, roosts, and detections during the breeding season for each species, complete spatial records for locations of surveys that resulted in non-detections were not available, which precluded our ability to reconstruct accurate detection–non-detection data sets. Therefore, we were limited to using a presence-only approach for these species, and we used the program MAXENT (Phillips et al. 2006) to develop territory distribution models.
We first evaluated whether these data sets met the assumptions critical for valid inference from a presence-only approach: (1) sampling is either random or representative throughout a landscape, and (2) detection probability is constant across sites (Yackulic et al. 2013). From 1987 to 2017, standardized surveys were conducted throughout the range of each species in the Lake Tahoe Basin, with no geographic bias; therefore, they provided a representative sample of the landscape. Although the survey protocols used over this 30-year period varied, they typically included multiple survey stations and multiple visits to those stations during the breeding season in a given year. These historical protocols typically yielded consistently high probabilities of detection (e.g., Woodbridge and Hargis 2006, Tempel and Gutiérrez 2013). Follow-up protocols after detection for each species provided additional data on the locations of important features of the species’ territories, nests, and breeding season roosts. Both the owl and the goshawk are relatively long-lived taxa with high fidelity to territories, and both attributes increase the likelihood of detecting territories over multiple years of surveys.
MAXENT can estimate species occurrence probability if the assumptions listed above are met (which they were), and users have knowledge of the occurrence probability of a species under “average” conditions (Phillips and Dudik 2008). To explicitly determine occurrence probability for each species, first we used the logistic model output to define the minimum probability thresholds for the occurrence of known territory centers and did not use the default threshold of 0.5. Second, to provide more realistic estimates of the proportion of territories that are actually occupied above the minimum occurrence probability thresholds, we evaluated how variation in occurrence probability across entire territory areas affected actual territory occupancy. To evaluate this relationship, we calculated the cumulative occurrence probability by summing occurrence probability across all pixels in each territory and compared them to the proportion of territories actually occupied across multiple cumulative probability bins. We predicted that above the minimum threshold of cumulative summed occurrence probability, (1) the proportion of potential territories actually occupied should also increase as the cumulative occurrence probability increases, and (2) not all sites above the minimum cumulative threshold for territory occurrence would be occupied. The minimum cumulative occurrence probability threshold and proportional relationships between cumulative occurrence probability and true occurrence were both used to estimate the number of suitable territories likely to be occupied in future decades.
Available data for the owl and goshawk included breeding season detections, roost locations, and nest locations during the nesting period that had been assigned to geographic areas and identified as territories by local agency biologists. There were 20 known and two suspected California Spotted Owl territories from 1998 to 2017. Nineteen of the 22 territories (86%) were occupied within 5 years of the baseline vegetation year for the simulations (2010), and 10 of the 22 (45%) had nest locations. There were 28 Northern Goshawk known territories from 1992 to 2017. Twenty-six of the 28 (93%) territories were occupied within 5 years of 2010, and 26 of the 28 (93%) had nest locations. Both the owl and goshawk are considered to be central-place foragers that concentrate activities around nests and roosts, and foraging activity declines with increasing distance from nests or roosts (Carey et al. 1992, Ward et al. 1998). To select a focal point for analysis of each territory, we used the following criteria: (1) the location of the active nest closest to the year 2010, and (2) if there was no known nest, the centroid of nesting season detections. For goshawk territory centers, 93% used nest locations and 7% used nesting season detection centroids. For the owl territory centers, 45% used nest locations and 55% used nesting season detection centroids.
For the Pacific marten, occupancy surveys had been conducted throughout the Lake Tahoe Basin, but they are not capable of reliably identifying the sex of an individual or the breeding status of females, which are necessary for identifying territories that support reproduction. From 2009 to 2011, demographic characteristics and spatial locations of marten were studied in five study areas that included the variation in mesic to xeric forest types and elevation ranges in which martens in the Lake Tahoe Basin occur (Slauson 2017). Although the five study areas were not distributed throughout the Lake Tahoe Basin, they provided a representative sample of the range of conditions in which martens occur. Territory centers for martens in this study were estimated using a spatially explicit capture–recapture approach and a capture protocol that yielded capture probabilities > 0.90 (Slauson 2017). While there were options for developing a marten model with presence–absence data, we chose the presence-only approach to be consistent with that used for the two other old-forest-associated predators. Like the owl and goshawk, female martens become central-place foragers during the denning period, where their activities are concentrated near their dens (e.g., Henry et al. 1997), which makes territory centers estimated during the denning period appropriate for developing models of territory occurrence. We used estimated territory centers for 16 female martens studied from 2009 to 2011 (Slauson 2017) to develop the marten territory distribution model.
Availability data sets (background points) for each old-forest-associated predator were developed by generating random points that matched the spatial extent of each predator’s presence data throughout the Lake Tahoe Basin (owl, goshawk) or that were limited to the geographic extents of the five study areas on the west and south shore of the Lake Tahoe Basin (marten). The number of random points used for each predator’s availability data set was scaled so the presence data would represent a prevalence of > 10%, which resulted in 250 points for the goshawk, 200 for the owl, and 150 for female martens (van Proosdij et al. 2016).
Candidate predictor variables
We used a combination of reviews of published literature on each species and personal experience with each taxon to identify candidate variables. We also considered LANDIS-II output variables (e.g., biomass of woody vegetation) that were potential analogs to variables known to be influential in old-forest-associated predator occurrence or could be alternatives over standard variables (e.g., California Wildlife Habitat Relationships tree size classes). For the goshawk, we selected a subset of the variables that have been shown to be influential in developing a goshawk habitat suitability model for the Sierra Nevada-Cascade bioregion (Dunk et al. 2012). For the owl, we used a recent review and synthesis of spotted owl habitat relationships (Gutiérrez et al. 2017) to identify candidate variables. For the marten, we relied primarily on the prior analysis of Slauson (2017) for habitat relationships with marten density during the denning season to identify candidate variables. Appendix 1 provides a complete list and description of all candidate variables; Appendix 2 shows the process of developing candidate variables from LANDIS-II variables.
Selection of spatial scales for each variable
In the development of territory distribution models for each old-forest-associated predator, we evaluated each variable at spatial scales representing core areas up to the entire territory because multiscale models often outperform models that measure resources at a single scale (McGarigal et al. 2016). We defined “core areas” as the areas of concentrated use that contain combinations of roosting, nesting, and foraging (owls [Blakesley et al. 2005, Williams et al. 2011] and goshawk [reviewed in Woodbridge and Hargis 2006]) or denning and foraging (female martens [Slauson 2017, Slauson and Zielinski 2019]) that can influence survival and reproduction. To select the optimal scale for each variable, we evaluated statistical fit of each scale at which a variable was measured (Shirk et al. 2012). Each candidate resource covariate was measured at four to six spatial scales around each territory’s focal point (Table 1). Variables were measured at each spatial scale with circular moving windows using the FOCALSUM or FOCALMEAN functions in ArcGIS (ESRI, Redlands CA, 92373). Spatial scales for each variable were compared between old-forest-associated predator territories and available points using t tests, and the scale with the lowest P value was retained for model development (Shirk et al. 2012). The final step in evaluating candidate variables was to conduct a correlation analysis, and when a pair of variables had an r2 > 0.6, the variable selected for exclusion was based on the following criteria: (1) remove the variable with the higher P value, and (2) remove the variable that is highly correlated with the most other variables. The final candidate variable set for each old-forest-associated predator included a set of uncorrelated variables measured at their optimal spatial scale.
Estimating the number of occupied territories through time
To apply the old-forest-associated predator territory distribution models to estimating the number of territories through the 100-year modeling time frame, the following questions were addressed for each species: (1) what is the minimum distance between territories? (2) what is the minimum occurrence probability threshold for territory occupancy? and (3) how does the proportion of suitable territories occupied change as the cumulative occurrence probability increases?
California Spotted Owl
The 22 owl territories were buffered with radii ranging from 625 to 1950 m. The buffer distance of 1200 m optimized minimizing the amount of overlap with adjacent territories and included the largest core area, and was selected as the minimum territory distance. This distance is similar to the average nearest-neighbor distances (1100 m) reported by researchers at lower elevation sites (Seamans and Gutiérrrez 2007, Tempel et al. 2014).
Northern Goshawk
Where forest habitats are continuous, spacing between nests of goshawks is fairly regular (Woodbridge and Hargis 2006). In the Klamath and Modoc National Forests in California, the mean nearest-neighbor distance between 59 nesting pairs in the Klamath was 3.3 km (SE = 0.3) (Dietrich and Woodbridge 1994); similar spacing was observed in the Modoc (Woodbridge and Hargis 2006). Therefore, buffers equivalent to 50% of the nearest-neighbor distance (1.65 km) were used.
Pacific marten
In California, home ranges of female Pacific and Humboldt martens average ~300 ha (Spencer et al. 1983, Slauson and Zielinski 2019), and individual females have never been detected at adjacent sample units when a 2-km systematic grid has been used to monitor a population of Humboldt martens (K. Slauson, pers. obs.), which further suggests that 300 ha or 1-km radius circles represent a reasonable average territory size for female martens in California. Therefore, buffers equivalent to an area of 314 ha, 1000-m radius was used.
RESULTS
Model development and evaluation
California Spotted Owl
There were 18 candidate variables, with 13 pairs of highly correlated (r2 > 0.6) variables. Ten of the highly correlated variables were removed (Table 2). The MAXENT model for owl territory distribution was developed from a final candidate set of eight variables (Table 2). Of these eight variables, only five were highly influential in differentiating between territory centers and random points (Table 2). Seven of the eight variables were used in the final model, which had a presence-only area under the curve (AUC) = 0.88. The threshold at which the sum of the sensitivity (true positive rate) and specificity (true negative rate) is highest was 0.57, and that was used as the final probability threshold for identifying suitable and unsuitable owl territory centers. The use of this probability threshold resulted in correctly classifying 86% (19/22) of spotted owl territory centers.
The five most influential variables accounted for 94% of the relative contribution of all variables in the spotted owl territory distribution model: elevation (30%), total biomass in the highest range (> 10,549 kg/ha [24%]), trees > 200 years of age (19%), annual precipitation (15%), and urban development (8%) (Table 2, Fig. 1). In the Lake Tahoe Basin, California Spotted Owls are at their upper elevational limit, and home range cores had a mean elevation of 2151 m, with a range from 1913 m to 2394 m. High total biomass was most influential at the territory scale (1600 m) and composed an average of 27% (217 ha) of spotted owl territories. Annual precipitation was higher at the territory scale (2350 m) than at available points, which reflected the decline in owl territories distributed from the mesic west to xeric east sides of the Lake Tahoe Basin. The presence of one or more trees > 200 years of age was most influential at the core-area scale (625 m), and while it represented only an average of 3.5% (4.3 ha) of the core area, it was most likely related to the presence of large old trees with features suitable for nesting and roosting.
Northern Goshawk
There were 19 candidate variables, with five pairs of highly correlated variables; five variables were removed (Table 2). Of the 14 candidate variables, only five were highly influential in differentiating between goshawk territory centers and available points. Seven of 14 variables were used in the final model, which had a presence-only AUC = 0.90. The threshold at which the sum of sensitivity and specificity was highest was 0.61; however, this threshold had a lower sensitivity (70%) for territory centers than desired. The threshold of 0.3 increased the sensitivity, correctly classifying 89% (25/28) of territory centers.
The five most influential variables in the goshawk model accounted for 89% of the relative contribution of all variables to the final model: slope (25%), trees 80–150 years old (21%), poor habitats (20%), urban development (13%), and total biomass of woody vegetation in the highest class (10%) (Table 2, Fig. 1). Core areas (500 m) of territories were on more gentle slopes and were composed of a greater amount (48%) of forest with trees aged 80–150 years than was available in the Lake Tahoe Basin (Table 2). Core areas of territories included less urban habitat than was available, and at the territory scale (2350 m), included less poor habitat than was available in the Lake Tahoe Basin. Core areas also had more forest in the high biomass category than was available (Table 2).
Pacific marten
There were 20 candidate variables, with five pairs of highly correlated variables; five correlated variables were removed (Table 2). Of the 15 candidate predictors used to develop the female marten model, only four had > 5% relative contribution to the final model (Table 2), which had a presence-only AUC = 0.96. The threshold at which kappa was optimized was 0.96; however, that threshold had a lower sensitivity (53%) than desired. The threshold of 0.43 increased sensitivity and correctly classified 88% (14/16) of all female territory centers and 93% (13/14) of territory centers for denning adult females. The range of 0.19–0.43 was used to identify low-suitability female territory centers, and accounted for the two sub-adult females’ and one adult female’s territory center.
The four most influential variables accounted for 88% of the relative contribution of all variables to the final model: mesic late seral forest with moderate to closed canopies (53%), presence of trees > 180 years of age (17%), mesic mid seral forest with moderate to closed canopies (9%), and poor habitats (9%) (Table 2, Fig. 1). Mesic late seral forest was most influential at the core-area scale (250 m), composing an average of 57% of the 20-ha cores of female territories. Trees > 180 years old were most influential at the territory scale (1200 m), where territories had an average of 36.1 ha with trees in this age class present.
Estimating the number of occupied territories
California Spotted Owl
Using the 1200-m buffers, occupied owl territories had a minimum cumulative occurrence probability threshold sum of 152.4. Above that threshold, the proportion of available suitable territories that were actually occupied was not equally distributed across cumulative occurrence probability bins and increased from 22%, for probability sums ranging from 154 to 225, to 56% as the probability sum became > 225 (Appendix 3). Therefore, these bin-specific territory occurrence rates were used to estimate the proportion of suitable territories likely to be occupied in any future time step.
Northern Goshawk
Using the 1650-m buffers, occupied goshawk territories had a lower threshold of the sum of occurrence probability of 44.5; however, 85% (22 of 26) of territories had sums > 100, and the mean sum of occurrence probability of all 26 territories was 273.9 (SE = 33). Above the sum of 100, the proportion of occupied territories was not equally distributed across cumulative probability bins and increased from 24% (100–200) to 64% as cumulative occurrence probability became > 200 (Appendix 3). Therefore, these bin-specific occurrence rates were used to estimate the proportion of territories likely to be occupied in any future time step.
Pacific marten
Using the 1000-m buffers, female marten territories had a lower threshold of the sum of occurrence probability of 84.5; however, 69% (11 of 16) of territories were > 150, and the mean sum of habitat suitability of all 16 territories was 173.0 (SE = 16). Above the lower threshold, the proportion of occupied territories was not equally distributed across probability bins and increased from 13% (< 100) to 19% (100–150) to 69% as the sum of occurrence probability became > 150 (Appendix 3). These bin-specific territory occupancy rates were used to approximate the proportion of territories likely to be occupied by female marten in future decades.
Changes in landscape composition from growth, wildfire, and thinning
At the start of the simulation period (2010), the Lake Tahoe Basin was composed of ≤ 10% late seral forest that contained the characteristics most important for supporting territories of all three old-forest-associated predators (Fig. 2, 3). By the end of the simulation period, the portion of the landscape’s composition of late seral forest increased 3–5 times under all five scenarios (Appendix 3). Wildfire, both managed and unmanaged, burned approximately 9% of the Lake Tahoe Basin per decade, with high-severity fire composing approximately 1–2% per decade. Wildfire frequency peaked from 80 to 100 years with approximately 25% of the Lake Tahoe Basin burned, and that was 2–9% greater than any prior two decades. Thinning treatments occurred in an average of 17.3% (Scenario 2), 43.1% (Scenario 3), 14.5% (Scenario 4), and ~15.0% (Scenario 5) of owl-sized territories across the Lake Tahoe Basin, and mechanical thinning comprised only 23–28% of thinning treatments. Due to their low-elevation distribution, known owl territories had a much higher degree of overlap with both WUI zones than did goshawk or marten territories; consequentially, known territories had 53–82% more thinning treatments than the overall landscape, with nearly all of that increase due to hand thinning treatments. By the end of the simulation period, known owl territories had cumulative thinning totals of 238% (Scenario 2), 592% (Scenario 3), 155% (Scenario 4), and 155% (Scenario 5), which suggests that known territories will be treated 1.5–5.9 times over the next 100 years (Fig. 3).
Trends in estimated number of occupied territories
California Spotted Owl
The estimated number of occupied owl territories exhibited short-term increases of > 50% for Scenarios 1, 2, and 4 over the first 20–40 years, followed by slow declines out to 80 years (Fig. 3). Although the short-term increase in the estimated number of occupied territories was slower for Scenario 3, the period of increase extended over the first 80 years when all other scenarios had started to decline (Fig. 3). Scenario 5 had the earliest (20 years) and lowest peak in the increase in occupied territories and the fewest number of occupied territories by the end of the simulation period (80–100 years) (Fig. 3). Beyond 80 years, all scenarios showed the sharpest decline after the period with the most wildfire. All scenarios maintained a higher number of occupied territories than at the start, except Scenario 5, but Scenarios 1, 2, and 4 performed better than Scenarios 3 and 5 using this criterion alone (Appendix 4). The cumulative amount of high-severity wildfire over 100 years under Scenarios 3 and 5 in occupied owl territories (20.9%) and potential owl territories (11.5%) was 5–19% lower than for Scenarios 1, 2, and 4 (Fig. 3).
Northern Goshawk
The estimated number of occupied goshawk territories exhibited slow increases of 20–33% over the 100-year simulation period for Scenarios 1–4 (Fig. 3). Scenario 5 had a much more rapid short-term (20 years) and greater overall (200%) increase in occupied territories. Scenarios 1–4 ended up with increases in the number of territories of 5.1–7.3 over the start in 2010, with only small (≤ 2.2 territories) differences between scenarios (Fig. 3, Appendix 4). Scenario 5 clearly outperformed the other four scenarios and supported a doubling of the number of occupied territories over the simulation period.
Pacific marten
The estimated number of occupied female marten territories exhibited rapid increases of 50% over the first 40 years across all five scenarios (Fig. 3). Each scenario ended up with increases in the number of territories of between 19 and 52 territories across the Lake Tahoe Basin over the estimated number of occupied territories at the start of the simulations, with Scenario 5 performing poorest overall (Fig. 3, Appendix 4).
DISCUSSION
The strength of our conclusions about the responses of the old-forest-associated predators to the five simulated management scenarios is entirely dependent on how accurately each model captured the features that are important to territory occupancy and how accurately they predicted the effects of changes in these features from forest growth and disturbance. Our goal for developing the empirical models using local data sets from the Lake Tahoe Basin was to create models that reflected the specific conditions in the Lake Tahoe Basin and provided greater accuracy than using models developed elsewhere or for larger geographical scales. However, a limitation of presence-only modeling is that currently there are no diagnostics for quantitatively assessing model fit and accuracy, and although AUC is often used, it has methodological shortcomings (Yuckulic et al. 2013). Therefore, we relied on critically evaluating modeled relationships and their consistency with published literature for each old-forest-associated predator to evaluate our models.
For the California Spotted Owl, two important biophysical variables—elevation and annual precipitation— accurately reflected the known elevational limitations of its distribution in the Lake Tahoe Basin (Gutiérrez et al. 2017) and limited territory establishment on the more mesic western half of the Lake Tahoe Basin over the last 30 years. At the territory scale, the positive relationship with increasing amounts of forest habitat in the high total biomass category, the category with the highest compositions of biomass in trees >100 years old (Appendix 2), is consistent with studies throughout the owl’s range that have shown that territories contain a greater proportion of mature forest with large trees than is available (reviewed in Roberts 2017). At the core-area scale, although owl territory centers had higher mean amounts of forest habitat with trees > 200 years, the response curve suggests a negative relationship as this age class increases. This result is contrary to several studies that have shown that owl occupancy, adult survival, and reproductive success are positively associated with the proportion of the core area containing structurally complex conifer forest with large trees and high canopy cover (Blakesley et al. 2005, Seamans and Gutiérrez 2007, Tempel et al. 2014). Only 39% of owl territories had forest with trees > 200 years of age in 2010, and when it was present, it was a small amount (3.5%) of the core area, suggesting a modeled relationship that is of limited biological significance and is likely a limitation on the accuracy of the model. The most significant relationship observed at the core-area scale in regard to tree age classes was for forest habitat with trees 80–160 years old, which was significantly greater in owl core areas (1200 m, X̅ = 51 ha [42% or core]) compared to that which was available (X̅ = 35 ha [29%]; t = -3.1, P = 0.001); although consistent with the literature, this variable was excluded from the analysis because it was strongly positively correlated with biomass.
The goshawk territory centers were most influenced by variables measured at the core-area scales (500–700 m); the importance of this scale in goshawk territory placement has been shown by a number of studies in North America (reviewed in Hansen 2012). Nest sites in California are often on gentle slopes (reviewed in Hansen 2012), and in the Lake Tahoe Basin territory core areas, which encompass the nest sites, the sites were also located on gentle slopes. Core areas in several North American studies have also been shown to contain more mature forest (reviewed in Hansen 2012), and our findings of a positive relationship between occurrence probability and increasing amount of forest habitat with trees 80–150 years old is consistent with that pattern. Goshawk territories in the Lake Tahoe Basin exhibited negative responses to urban development at the core-area scale, which is consistent with other studies that have compared proximities of nests to human development (e.g., Bosakowski and Speiser 1994). At the territory scale, there was a negative relationship to poor habitats, which suggests that territory centers are located in areas with a minimal amount of mature mid- to low-elevation forest. This relationship is also consistent with most studies of goshawks in California (reviewed in Hansen 2012) and with the habitat associations of many of the goshawk’s most important prey in the Lake Tahoe Basin (Keane 1999).
The female Pacific marten territory model had the single most influential variable across all three old-forest-associated predators, with 53% explained by the amount of mesic late seral forest with moderate to closed tree canopy at the core-area scale. This same variable best explained variation in female density, using spatially explicit capture–recapture analysis and the same data set (Slauson 2017), and is consistent with habitat selection for older forest conditions by females elsewhere in the Sierra Nevada (e.g., Spencer et al. 1983). The positive step functions for trees > 180 years old at the territory scale suggest that a minimal threshold for the presence of patches of late seral forest distributed throughout territories is important and is consistent with female martens moving their kits to other parts of their home range where suitable den structures occur (e.g., Nichols 2016), and with their need for suitable rest structures in large-diameter live and dead trees year-round (e.g., Spencer et al. 1983). The positive step function for a minimum amount of mesic mid seral forest in combination with late seral forest in the territory core area is also consistent with recent analyses of female marten home range compositions elsewhere in California (K. Slauson, pers. comm.). Finally, the negative relationship with poor habitats is consistent with Thompson et al.’s (2012) review that marten home ranges are most often composed of ~70% suitable habitat.
Overall, each old-forest-associated predator’s territory distribution model included variables and modeled relationships that were either all (goshawk, marten) or nearly all (owl) consistent with the published literature and also captured important local relationships to the Lake Tahoe Basin. These models should therefore provide reasonable approximations of the main factors influencing territory distribution. Another strength of our approach for the old-forest-associated predator models was the use of the relationship between the proportion of the territories actually occupied and how much of the territory was composed of suitable habitat. This relationship followed our prediction that as the proportion of the territory increased in suitable habitat, so too did the proportion of the territories actually occupied. This allowed us to scale our estimates of the actual number of territories that would be occupied using these relationships. This assumes that these relationships are likely to remain the same from 2010 to 2110, and if so, should provide more accurate predictions.
One important caveat about our models is that they assume that habitat features that were important in 2010 will continue to be as important in the future. While our review of each influential variable suggests most are consistent with the literature pertinent to each old-forest-associated predator, they cannot account for novel conditions produced from natural or anthropogenic disturbances that may also be suitable for the species. For example, if thinning treatments from below create suitable conditions with larger diameter trees but in lower total biomass ranges than we found influential for spotted owls, our model will not be able to account for such novel conditions. In addition, while we feel the old-forest-associated predator data sets met the two key assumptions for using a presence-only modeling approach, if the assumption of consistent probability of detection over 30 years of survey effort was not met, it could result in biases in the modeled relationships between territory occurrences and habitat variables that reduce the accuracy of their responses to the simulated landscape changes.
The overriding factor that caused increases in the number of occupied territories for all three old-forest-associated predators across all five management scenarios was the increase in the recruitment of late seral forest from growth and succession in the Lake Tahoe Basin over the 100-year simulation period. Each predator’s model for territory occurrence probability had variables associated with late seral forest conditions among the most influential variables that could change during the simulations. The recruitment of late seral forest far outpaced the loss of this seral stage from simulated natural disturbances that include high-severity wildfire or bark beetle outbreaks. This result highlights the importance that the existing landscape conditions and their forest growth trajectories can have on minimizing the risks of negative impacts from management alternatives similar to those simulated in our study. We would not expect these same results had the Lake Tahoe Basin been composed of a greater mosaic of age classes or a greater mosaic of forest and non-forest habitat.
With the exception of Scenario 5 for goshawk, there were only relatively minor differences in numbers of territories estimated to be occupied between scenarios across the three old-forest-associated predators. Scenario 5 performed best for goshawk and poorest for the owl and female martens. The pace and scale of the combination of thinning and fire was greatest for Scenario 5, and although it resulted in the most rapid increase in late seral forest across the Lake Tahoe Basin, it also had the largest reduction in patch sizes of late seral forest compared to the other scenarios. The territory suitability models for owl and female marten both had influential variables associated with late seral forest at the territory scale, while the model for goshawk did not. This difference in the influence of spatial scales between the goshawk model (core scale only) and owl and marten (core and territory scales) appears to explain this difference in response to Scenario 5.
The second largest difference in numbers of territories estimated to be occupied between scenarios was for Scenario 3 for the California Spotted Owl, which although having fewer total occupied territories at any decade, had the most sustained increase compared to all other scenarios. This period of increase continued for 40 years (years 40–80) when all other scenarios showed a decline in the number of occupied territories. This suggests that Scenario 3 created forest habitat that is less susceptible to high-severity wildfire; therefore, territories were more resilient to change from this type of disturbance. Scenario 3 had the most thinning, including both mechanical and hand, and the prescriptions used removed more total biomass, which appears to reduce the relative rate of territory increase compared to the other scenarios. However, toward the latter half of the century, Scenario 3 was the only one in which the numbers of occupied owl territories increased, which suggests a trade-off between short-term habitat degradation shifting to longer term gains of creating more resilient forest conditions to uncharacteristically severe wildfire.
Scenario 3 limited the cumulative percent of owl territories burned by high-severity wildfire over 100 years to < 20% compared to 25–40% for the other scenarios (Fig. 3). The benefits of Scenario 3 for owls may continue to accumulate beyond 100 years relative to the other scenarios. While Scenario 3’s higher pace and scale of fuels treatments reduced the loss of suitable habitat from severe wildfire, those benefits appeared to not increase after 50 years relative to the other scenarios (Fig. 3). This suggests that an aggressive short-term management alternative may put the landscape on the right trajectory to being more resilient to severe wildfire but that reducing the pace and scale after 50 years may be warranted. However, Scenario 5 appeared to have crossed the threshold of pace and scale for the owl and marten such that short-term negative effects outweighed the long-term benefits of reducing the risk of severe wildlife and increasing landscape resiliency. The increase in the pace and scale of treatments in Scenario 5 appeared to negatively affect the landscape composition for owls and female martens by reducing patch sizes of late seral forest that are important to both species at their territory scales. Overall, trends in territory numbers for all management scenarios were positive for the first half of the century, which suggests that management approaches in the ranges of Scenario 3 and 4 may result in a balance between moving the Lake Tahoe Basin landscape more rapidly toward resiliency and increasing the numbers of territories for all three old-forest-associated predators. However, trends toward the latter half of the century start to become negative for the owl across all scenarios, which suggests that the selection of management strategy from that point may need to be revisited, especially in the limited areas where the owls occur in the Lake Tahoe Basin. Of all three old-forest-associated predators, the owl’s distribution in the Lake Tahoe Basin had the largest proportional overlap with the management zones (WUIs) that received the most treatments across Scenarios 2–5, which suggests that over the long-term, treatments may cross a threshold where they are no longer beneficial in maintaining habitat suitability by reducing the risk of severe wildfire, but rather become detrimental.
Our simulation results are consistent with the relative differences between the short-term effects of mechanical treatments (worse; Stephens et al. 2014, Tempel et al. 2014) versus wildfire (better; Roberts et al. 2011, Lee and Bond 2015) treatments in California Spotted Owl habitat that have been demonstrated elsewhere in field studies. However, for marten, mechanical treatments appeared to perform slightly better than mechanical treatments with wildfire. We caution against interpreting our results and making strong conclusions about the relative effects of these two fuels treatment alternatives because our models could assess only the relative effects of their changes in coarse stand characteristics, which does not capture how changes in forest structure in the understory can influence old-forest-associated predator foraging behavior and prey abundance. In general, each predator exhibited more adverse responses to mechanical fuels treatments than to prescribed fire in field studies, even when the thinning was limited to understory trees and shrubs only (e.g., Stephens et al. 2014). Treatments that mimic the heterogeneity resulting from the natural process of wildfire and result in diverse forest structures and large patches with old-forest characteristics will be the most successful in achieving objectives for both fuels reduction and suitable habitat conservation for old-forest-associated predators.
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.ACKNOWLEDGMENTS
We would like to thank Stephanie Coppeto and Shay Zanetti at the Lake Tahoe Basin Management Unit and Daniel Shaw at California Department of Parks and Recreation for providing data on owl and goshawk territories. The Southern Nevada Public Land Management Act provided funding for this project.
DATA AVAILABILITY
The data/code that support the findings of this study are available upon request from the corresponding author, KS. None of the data/code are publicly available because they contain specific breeding locations for several sensitive wildlife species. Ethical approval for the marten data used in this study was provided by the California Department of Fish and Wildlife (SC-4683) and Nevada Department of Wildlife (S-31799).
LITERATURE CITED
Agee, J. K. 1993. Fire ecology of Pacific Northwest forests. Island Press, Washington, D.C., USA.
Blakesley, J. A., B. R. Noon, and D. R. Anderson. 2005. Site occupancy, apparent survival, and reproduction of California Spotted Owls in relation to forest stand characteristics. Journal of Wildlife Management 69:1554-1564. https://doi.org/10.2193/0022-541X(2005)69[1554:SOASAR]2.0.CO;2
Bosakowski, T., and R. Speiser. 1994. Macrohabitat selection by nesting Northern Goshawks: implications for managing eastern forests. Studies in Avian Biology 16:46-49.
Carey, A. B., S. P. Horton, and B. L. Biswell. 1992. Northern Spotted Owls: influence of prey base and landscape character. Ecological Monographs 62:223-250. https://doi.org/10.2307/2937094
Collins, B. M., S. L. Stephens, G.B. Roller, and J. J. Battles. 2011. Simulating fire and forest dynamics for a landscape fuel treatment project in the Sierra Nevada. Forest Science 57:77-88.
Crocker-Bedford, D. C. 1990. Goshawk reproduction and forest management. Wildlife Society Bulletin 18: 262-269.
Dietrich, P. J., and B. Woodbridge. 1994. Territory fidelity, mate fidelity, and movements of color-marked Northern Goshawks in the Southern Cascades of California. Studies in Avian Biology 16:130-132.
Dunk, J., N. Nichol, B. Woodbridge, D. L. Hansen, and D. L. Laplante. 2012. Range, distribution, and abundance. Pages 43–77 in B. Woodbridge, D. L. Hanse, and J. R. Dunk, editors. The Northern Goshawk in California: a technical assessment of its current status and ecology. Report to Region 5 of the U.S. Forest Service.
Gutiérrez, R. J., P. N. Manley, and P. A. Stine. 2017. The California Spotted Owl: current state of knowledge. General Technical Report PSW-GTR-254. U.S. Forest Service, Albany, California, USA. https://doi.org/10.2737/PSW-GTR-254
Hansen, D. L. 2012. Nesting habitat. Pages 128–157 in B. Woodbridge, D. L. Hanse, and J. R. Dunk, editors. The Northern Goshawk in California: A technical assessment of its current status and ecology. Report to Region 5 of the U.S. Forest Service.
Henry, S. E., E. C. O’Doherty, L. F. Ruggiero, and W. D. Van Sickle. 1997. Maternal den attendance patterns of female American marten. Pages 78-85 in G. Proulx, H. N. Bryant, and P. M. Woodward, editors. Martes: taxonomy, ecology, techniques, and management. Provincial Museum of Alberta, Edmonton, Alberta, Canada.
Keane, J. J. 1999. Ecology of the Northern Goshawk in the Sierra Nevada, California. Dissertation, University of California, Davis, California, USA.
Lee, D. E., and M. L. Bond. 2015. Occupancy of California Spotted Owl sites following a large fire in the Sierra Nevada, California. Condor 117:228-236. https://doi.org/10.1650/CONDOR-14-155.1
Loudermilk, E. L., R. M. Scheller, P. J. Weisberg, J. Yang, T. E. Dilts, S. L. Karam, and C. Skinner. 2013. Carbon dynamics in the future forest: the importance of long‐term successional legacy and climate-fire interactions. Global Change Biology 19:3502-3515. https://doi.org/10.1111/gcb.12310
Maxwell, C., R. M. Scheller, J. W. Long, and P. Manley. 2022. Forest management under uncertainty: the influence of management versus climate change and wildfire in the Lake Tahoe Basin, USA. Ecology and Society 27(2):15. https://doi.org/10.5751/ES-13278-270215
McGarigal, K., H. Y. Wan, K. A. Zeller, B. C. Timm, and S. A. Cushman. 2016. Multi-scale habitat selection modeling: a review and outlook. Landscape Ecology 31:1161-1175. https://doi.org/10.1007/s10980-016-0374-x
Miller, J. D., H. D. Safford, M. Crimmins, and A. E. Thode. 2009. Quantitative evidence for increasing forest fire severity in the Sierra Nevada and southern Cascade Mountains, California and Nevada, USA. Ecosystems 12:16-32. https://doi.org/10.1007/s10021-008-9201-9
Moriarty, K. M., C. W. Epps, and W. J. Zielinski. 2016. Forest thinning changes movement patterns and habitat use by Pacific marten. Journal of Wildlife Management 80:621-633. https://doi.org/10.1002/jwmg.1060
Nichols, M. J. 2016. American marten denning behavior in Michigan. Thesis. Grand Valley State University, Allendale, Michigan, USA. http://scholarworks.gvsu.edu/theses/794
Phillips, S. J., R. P. Anderson, and R. E. Schapire. 2006. Maximum entropy modeling of species geographic distributions. Ecological Modelling 190:231-259. https://doi.org/10.1016/j.ecolmodel.2005.03.026
Phillips, S. J., and M. Dudík. 2008. Modeling of species distributions with Maxent: new extensions and a comprehensive evaluation. Ecography 31:161-175. https://doi.org/10.1111/j.0906-7590.2008.5203.x
Powell, R. A. 1994. Structure and spacing of Martes populations. Pages 101–121 in K. B. Aubry, W. J. Zielinski, M. G. Raphael, G. Proulx, and S. W. Buskirk, editors. Martens, sables, and fishers: biology and conservation. Cornell University Press.
Reynolds, R. T., R. T. Graham, M. H. Reiser, R. L. Bassett, P. L. Kennedy, D. A. Boyce, G. Goodwin, R. Smith, and E. L. Fisher. 1992. Management recommendations for the Northern Goshawk in the southwestern United States. General Technical Report RM-217, U.S. Forest Service, Fort Collins, Colorado, USA. https://doi.org/10.2737/RM-GTR-217
Roberts, S. L. 2017. California Spotted Owl habitat characteristics and use. Pages 49-73 in General Technical Report PSW-GTR-254, U.S. Forest Service, Albany, California, USA.
Roberts, S. L., J. W. van Wagtendonk, A. K. Miles, and D. A. Kelt. 2011. Effects of fire on spotted owl site occupancy in a late-successional forest. Biological Conservation 144:610-619. https://doi.org/10.1016/j.biocon.2010.11.002
Scheller, R. M., J. B. Domingo, B. R. Sturtevant, J. S. Williams, A. Rudy, E. J. Gustafson, and D. J. Mladenoff. 2007. Design, development, and application of LANDIS-II, a spatial landscape simulation model with flexible temporal and spatial resolution. Ecological Modelling 201:409-419. https://doi.org/10.1016/j.ecolmodel.2006.10.009
Scheller, R. M., A. Kretchun, T. J. Hawbacker, and P.D Henne. 2019. A landscape model of variable social-ecological fire regimes. Ecological Modelling 401:85-93. https://doi.org/10.1016/j.ecolmodel.2019.03.022
Scheller, R. M., W. D. Spencer, H. Rustigian-Romsos, A. D. Syphard, B. C. Ward, and J. R. Strittholt. 2011. Using stochastic simulation to evaluate competing risks of wildfires and fuels management on an isolated forest carnivore. Landscape Ecology 26:1491-1504. https://doi.org/10.1007/s10980-011-9663-6
Seamans, M. E., and R. J. Gutiérrez. 2007. Habitat selection in a changing environment: the relationship between habitat alteration and spotted owl territory occupancy and breeding dispersal. Condor 109:566-576. https://doi.org/10.1093/condor/109.3.566
Shirk, A. J., T. N. Wasserman, S. A. Cushman, and M. G. Raphael. 2012. Scale dependency of American marten (Martes americana) habitat relations. Pages 269-283 in K. B. Aubry, W. J. Zielinski, M. G. Raphael, G. Proulx, and S. W. Buskirk, editors. Martens, sables, and fishers: biology and conservation. Cornell University Press.
Slauson, K. M. 2017. Linking landscape pattern to population process in a carnivorous mammal. Dissertation, University of Montana, Missoula, Montana, USA.
Slauson, K. M., and W. J. Zielinski. 2019. Diets of denning female Pacific martens vary with the developmental stage of their kits. Ecology and Evolution 9:5963-5974. https://doi.org/10.1002/ece3.5179
Spencer, W. D., R. H. Barrett, and W. J. Zielinski. 1983. Marten habitat preferences in the northern Sierra Nevada. Journal of Wildlife Management 47:1181-1186. https://doi.org/10.2307/3808189
Stephens, S. L., S. W. Bigelow, R. D. Burnett, B. M. Collins, C. V. Gallagher, J. Keane, D. A. Kelt, M. P. North, L. J. Roberts, P. A. Stine, and D. H. Van Vuren. 2014. California Spotted Owl, songbird, and small mammal responses to landscape fuel treatments. BioScience 64:893-906. https://doi.org/10.1093/biosci/biu137
Sturtevant, B. R., E. J. Gustafson, W. Li, and H. S. He. 2004. Modeling biological disturbances in LANDIS: a module description and demonstration using spruce budworm. Ecological Modelling 180(1):153-174.
Syphard, A. D., R. M. Scheller, B. C. Ward, W. D. Spencer, and J. R. Strittholt. 2011. Simulating landscape-scale effects of fuels treatments in the Sierra Nevada, California, USA. International Journal of Wildland Fire 20:364-383. https://doi.org/10.1071/WF09125
Tempel, D. J., and R. J. Gutiérrez. 2013. Relation between occupancy and abundance for a territorial species, the California Spotted Owl. Conservation Biology 27:1087-1095. https://doi.org/10.1111/cobi.12074
Tempel, D. J., R. J. Gutiérrez, S. A. Whitmore, M. J. Reetz, R. E. Stoelting, W. J. Berigan, M. E. Seamans, and M. Z. Peery. 2014. Effects of forest management on California Spotted Owls: implications for reducing wildfire risk in fire‐prone forests. Ecological Applications 24:2089-2106. https://doi.org/10.1890/13-2192.1
Thompson, I. D., J. Fryxell, and D. J. Harrison. 2012. Improved insights into use of habitat by American martens. Pages 209-230 in K. B. Aubry, W. J. Zielinski, M. G. Raphael, G. Proulx, and S. W. Buskirk, editors. Biology and conservation of martens, sables, and fishers: a new synthesis. Cornell University Press, Ithaca, New York, USA.
United States Forest Service (USFS). 2004. Sierra Nevada forest plan amendment: final supplemental environmental impact statement. U.S. Forest Service, Vallejo, California, USA.
van Proosdij, A. S., M. S. Sosef, J. J. Wieringa, and N. Raes. 2016. Minimum required number of specimen records to develop accurate species distribution models. Ecography 39:542-552. https://doi.org/10.1111/ecog.01509
Ward Jr, J. P., R. J. Gutiérrez, and B. R. Noon. 1998. Habitat selection by Northern Spotted Owls: the consequences of prey selection and distribution. Condor 100:79-92. https://doi.org/10.2307/1369899
Westerling, A. L., and B. P. Bryant. 2008. Climate change and wildfire in California. Climatic Change 87:231-249. https://doi.org/10.1007/s10584-007-9363-z
Williams, P. J., R. J. Gutiérrez, and S. A. Whitmore. 2011. Home range and habitat selection of spotted owls in the central Sierra Nevada. Journal of Wildlife Management 75:333-343. https://doi.org/10.1002/jwmg.62
Woodbridge, B., D. L. Hansen, and J. R. Dunk, technical coordinators. 2012. The Northern Goshawk in California: a technical assessment of its ecology and status. Report to Region 5 of the USDA Forest Service.
Woodbridge, B., and C. D. Hargis. 2006. Northern Goshawk inventory and monitoring technical guide. General Technical Report WO-71. U.S. Forest Service, Washington, D.C., USA. https://doi.org/10.2737/WO-GTR-71
Yackulic, C. B., R. Chandler, E. F. Zipkin, J. A. Royle, J. D. Nichols, E. H. Campbell Grant, and S. Veran. 2013. Presence‐only modelling using MAXENT: when can we trust the inferences? Methods in Ecology and Evolution 4:236-243. https://doi.org/10.1111/2041-210x.12004
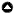
Fig. 1

Fig. 1. Variable response curves for the four most influential variables for MAXENT territory distribution models for the California Spotted Owl (Strix occidentalis occidentalis), Northern Goshawk (Accipiter gentilis), and female Pacific marten (Martes caurina) in the Lake Tahoe Basin of California and Nevada.
Fig. 2

Fig. 2. Baseline (2010) territory distribution maps for three old-forest-associated predators in the Lake Tahoe Basin, using existing territories and random availability points in a MAXENT species distribution modeling framework (A–C), and changes in the distribution and amount of territory occurrence probability 60 years beyond baseline (D–F).
Fig. 3

Fig. 3. Trends in the estimated number of occupied California Spotted Owl (Strix occidentalis occidentalis) (A–B), Northern Goshawk (Accipiter gentilis) (C–D), and female Pacific marten (Martes caurina) (E–F) territories for each management scenario from year 1 to 100 for the entire Lake Tahoe Basin and the Lake Tahoe West project area. Standard error bars were not included because they were all small, ranging from 0.2 to 1.4. G cumulative percent of known occupied spotted owl territories burned by high-severity wildfire over the 100-year simulation period.
Table 1
Table 1. Spatial scales of measuring candidate variables for modeling territory occurrence probability for the California spotted owl (Strix occidentalis occidentalis), Northern goshawk (Accipiter gentilis), and female Pacific marten (Martes caurina) in the Lake Tahoe Basin of California and Nevada.
| California Spotted Owl | Northern Goshawk | Pacific marten | |||||||
| Spatial scale | Area (ha) | Radius from nest/centroid | Area (ha) | Radius from nest/centroid | Area (ha) | Radius from home range center | |||
| Nest/den patches/ protected area center | 120 ha | 625 m | 80 ha | 500 m | 20 ha | 250 m | |||
| 80 ha | 500 m | ||||||||
| Reproductive core area | 200 ha | 800 m | 150 ha | 700 m | 175 ha | 750 m | |||
| 400 ha | 1200 m | 300 ha | 1000 m | ||||||
| Territory | 800 ha | 1600 m | 1130 ha | 1900 m | 300 ha | 1000 m | |||
| 1200 ha | 1950 m | 1750 ha | 2360 m | 450 ha | 1200 m | ||||
| 2000 ha | 3100 m | ||||||||
| References | Gutierrez et al. 2017 | Keane 1999 | Slauson 2017 | ||||||
| Roberts 2017 | Woodbridge et al. 2012 | Spencer et al. 1983 | |||||||
Table 2
Table 2. Mean values and mean percentage of territories represented by the most influential variables in the MAXENT model for California spotted owl (Strix occidentalis occidentalis), Northern goshawk (Accipiter gentilis), and female Pacific marten (Martes caurina) territory distribution in the Lake Tahoe Basin. Twenty-two spotted owl, 28 Northern goshawk, and 16 female Pacific marten territories were compared to 200, 250, and 150 random points matched to the spatial extent of each species dataset, respectively.
| Spatial scale | Means (ha/km) | ||||||||||||
| Variable | Radius from center | Area (ha) | MAXENT variable contrib. | Territory centers | Percentage of scale | Random points | |||||||
| California Spotted Owl | |||||||||||||
| Elevation | 800 m | 201 | 30.4% | 2156 m | NA | 2253 m | |||||||
| Total biomass highest | 1600 m | 804 | 22.7% | 217.0 ha | 27.0% | 149.2 ha | |||||||
| Tree max age > 200 | 625 m | 122 | 18.5% | 4.3 ha | 3.5% | 1.7 ha | |||||||
| Annual precipitation | 1950 m | 1194 | 14.6% | 982.6 | NA | 1021.4 ha | |||||||
| Urban development | 1950 m | 1194 | 8.0% | 106.4 ha | 8.9% | 219.9 ha | |||||||
| Northern Goshawk | |||||||||||||
| Slope | 500 m | 80 | 24.9% | 9.8% | NA | 14.4% | |||||||
| Tree max age 80–150 | 500 m | 80 | 20.8% | 38.0 ha | 47.5% | 22.4 ha | |||||||
| Poor habitats | 2350 m | 1750 | 20.2% | 614.2 ha | 35.1% | 872.3 ha | |||||||
| Urban development | 500 m | 80 | 12.8% | 1.2 ha | 1.5% | 8.9 ha | |||||||
| Total biomass highest | 700 m | 150 | 10.2% | 40.5 ha | 27.0% | 17.3 ha | |||||||
| Female Pacific marten | |||||||||||||
| Mesic late seral forest | 250 m | 20 | 52.8% | 11.4 ha | 57.0% | 6.1 ha | |||||||
| Tree max age > 180 | 1200 m | 450 | 17.0% | 36.1 ha | 8.0% | 19.7 ha | |||||||
| Mesic mid seral forest | 750 m | 175 | 9.1% | 50.2 ha | 28.7% | 32.7 ha | |||||||
| Poor habitats | 750 m | 175 | 8.9% | 74.3 ha | 42.5% | 83.8 ha | |||||||
| Variables excluded due to correlation > 0.6: Spotted owl-Maximum July temperature-1950 m, Road Density-1200 m, Deer mouse suitability-1950 m, Northern flying squirrel suitability-1600 m, Total biomass 10,549 to 31,668-1600 m, Trees 80-160 year old-1200 m; Northern goshawk-Stream density-3100 m, Maximum July temperature-700 m, Mesic mid and late seral forest with moderate to high canopy cover-700 m, Nesting season prey suitability-500 m, Trees < 50 years old-1900m; Female Pacific marten-Average May precipitation-250 m, Total woody biomass > 65,268-750 m, Minimum Jan temperature-750 m, Maximum July temperature-250 m. Variables contributing < 5% to final models: Spotted owl-Minimum Jan Temperature-800 m, Stream density-1600 m, Mean aspect-1950 m; Northern goshawk-Annual precipitation-500 m, Elevation squared-500 m, Minimum January temperature-700 m, All important prey suitability index-250 m, Minimum Jan temperature-700 m, Minimum April temperature-700 m, Trees > 190 years old-700 m, Mean aspect-500 m; Female Pacific marten-Total biomass 1335 to 3241 (kg/ha)-1200 m, Annual precipitation-500 m, Mean slope-250 m, All important prey suitability index-250 m, Minimum Jan temperature-750 m, Important denning season prey suitability index-250 m, Mean aspect-250 m, Stream density-750 m, Road density-250 m. | |||||||||||||