






The following is the established format for referencing this article:
White, A. M., T. G. Holland, E. S. Abelson, A. Kretchun, C. J. Maxwell, and R. M. Scheller. 2022. Simulating wildlife habitat dynamics over the next century to help inform best management strategies for biodiversity in the Lake Tahoe Basin, California. Ecology and Society 27(2):31.ABSTRACT
Many forests of the western United States have undergone over one hundred years of anthropogenic impacts that have led to increased tree density, homogenization in forest structure, and accumulation of woody material, which combined with a changing climate pose threats to valued social and ecological features. In California, recent waves of tree mortality and unprecedented large and destructive fires have led to rising concerns about the impact of these disturbances on biodiversity and how forest management actions can mitigate negative impacts. To better understand the degree to which different management scenarios could mitigate the negative impacts of these disturbances on biodiversity, we used a spatially explicit modeling platform to model forest management impacts on habitat for terrestrial vertebrate species in the Lake Tahoe Basin of California and Nevada. Specifically, we modeled how 5 different management scenarios that differed in the type of fuel reduction treatment (e.g., fire and mechanical removal of vegetation) and extent of area treated influenced the amount, value, and distribution of reproductive habitat for the 159 species present in the study area. Our model results suggested that within the study area forest growth was predicted to out-pace disturbance leading to a higher percentage of late seral conditions; however, choice of management strategy impacted the composition and structure of the forested landscape leading to different trajectories for wildlife. In general, scenarios that allowed for more extensive use of fire led to a more equitable distribution of habitat types, whereas extensive thinning by hand and mechanical methods resulted in future forest structure that provided better outcomes in terms of reproductive habitat for wildlife. Our modeling results also suggested that low to moderate management strategies were not likely to change the current trajectory to more dense forests dominated by fewer species.INTRODUCTION
In the seasonally dry forests of the western United States, fire is a natural disturbance that shapes habitat composition, structure, and distribution (Larson and Churchill 2012, Knapp et al. 2013, Lutz et al. 2018). In the mid-elevation forests of the Sierra Nevada Mountains in California, fire historically maintained a species-rich mixture of tree densities and size classes (Barbour et al. 2002, Beaty and Taylor 2007, North et al. 2009), but due to past practices such as logging, grazing, and highly effective fire suppression, mixed-conifer forests have become denser and simplified in structure and composition (Knapp et al. 2013, Stephens et al. 2013). Coupled with rapid changes in climate, these changes in forest structure have contributed to large-scale susceptibility of trees to bark beetle attacks during drought (Young et al. 2017) and massive, high-severity fires (Lydersen et al. 2014, Young et al. 2017). In California, theses large-scale disturbances threaten the State’s economy because these forests not only provide woody material, clean water, and abundant recreational opportunities, but also serve as habitat for a diverse community of plants and animals (Mayer and Laudenslayer 1988). The rapid change in these forests underscores the need to increase the pace and scale of forest treatments designed to reduce woody material and increase forest resiliency to future, large-scale disturbances (North et al. 2012, Stephens et al. 2013).
Forest treatments designed to reduce the risk of high-severity fire commonly focus on reducing the amount and connectivity of woody material on the forest floor (e.g., branches, logs, and shrubs), between the forest floor and the canopy (e.g., shrubs and small trees), and within the canopy (e.g., smaller, subdominant canopy trees; Agee and Skinner 2005). To implement large-scale forest restoration projects will require not only mechanical thinning of trees and shrubs, but due to the relatively small percentage of forest that can be mechanically treated because of logistical and fiscal constraints (North et al. 2015), will likely require the use of prescribed burning and managed wildfire (North et al. 2012). Although data collected from forested areas receiving both mechanical/hand removal of vegetation and prescribed burning treatments have been shown to have neutral to positive effects on many wildlife species (Fontaine and Kennedy 2012, Stephens et al. 2014), many species thrive in forests burning with moderate to high-severity fire effects (Smucker et al. 2005, Tingley et al. 2016, White et al. 2016), higher cover of shrubs (White et al. 2013), or complex, early seral habitat conditions (Swanson et al. 2011). Collectively, these studies confirm that wildlife response to fuel reduction treatments will depend on the type of disturbance or restoration action and on the resulting structure.
Forest structure, and its composition, provide the environmental conditions needed to support essential food, resting, and breeding requirements for wildlife. Changes to forest structure inherently impact the degree to which environmental conditions provide suitable habitat, i.e., improving habitat conditions for some species, while reducing it for others (Fontaine and Kennedy 2012, White et al. 2013, Stephens et al. 2014). The California Wildlife Habitat Relationships (CWHR) database (https://www.wildlife.ca.gov/Data/CWHR) classifies the suitability of habitat for vertebrate species based on predominate vegetation (tree, shrub, herbaceous, etc.), species composition, and stage of vegetation development (Fig. 1). For each combination of these conditions, CWHR provides a habitat suitability classification (high, moderate, low, unsuitable) for species based on expert-solicited information (Fig. 2A). These broad habitat classifications make it possible to predict habitat suitability for a rich diversity of wildlife species over large areas and timescales where data exist on vegetation type, structure, and seral stage.
We build on previous efforts to better understand how the type and intensity of management treatments are likely to influence wildlife habitat (e.g., Fontaine and Kennedy 2012, White et al. 2013, Stephens et al. 2014) over large spatial and temporal scales. Although providing for habitat connectivity and a mosaic of forest conditions provides general principles for landscape scale wildlife management, specific targets are rarely quantified and will differ with species and existing forest conditions (North and Manley 2012). Using a modeling framework, we assess how different types of forest restoration treatments (i.e., fire suppression, mechanical and hand thinning, prescribed burning, and managed wildfire) and placement, i.e., wildland-urban interface (WUI) or general forest, impact the quality and distribution of reproductive habitat for terrestrial vertebrate species in the Lake Tahoe Basin of California over the next century. We predicted that management scenarios incorporating managed wildfire would lead to greater structure complexity in forest types and subsequently provide greater benefits to wildlife than management scenarios focusing on mechanical treatments in the wildland-urban interface. This effort is part of a larger initiative led by the Lake Tahoe West Restoration Partnership to restore the social-ecological resilience of forests, watersheds, and communities on 59,000 acres of Lake Tahoe’s west shore (Abelson et al 2022).
METHODS
The Lake Tahoe Basin, located on the eastern crest of the Sierra Nevada Mountains straddling the states of California and Nevada, supports high levels of biodiversity (Fig. 1). The location of the basin between two major biogeographic provinces, the Sierra Nevada and the Great Basin, creates a juxtaposition of habitats, each province of which supports a unique array of species. In addition, the basin’s steep elevational gradients (ranging from 1900 m at Lake Tahoe’s surface to 3400 m at the highest mountain peak) span lower montane, upper montane, subalpine, and alpine habitats. Within these habitats, differences in forest structure and composition further influence the provisioning of resources used by wildlife for reproduction, foraging, and cover. Our assessment focused on the habitat of 159 vertebrate species that have suitable habitat for breeding in the Lake Tahoe Basin.
Dynamic modeling of habitat
To model habitat change over time, we used the landscape disturbance and succession model LANDIS-II (https://www.landis-ii.org/) to simulate forest change as a function of growth and succession (Mladenoff 2004, Scheller et al. 2007). Within LANDIS-II, trees and shrubs are modeled as species-age cohorts; each species has its own unique life history attributes and as such, each responds uniquely to disturbance. Every cell (1-ha resolution) across the landscape has a unique combination of cohorts, live biomass, dead material, and soils as determined by its simulated history of succession and disturbance type and intensity. In addition to fire and management, several species of bark beetles, including the Jeffrey pine beetle (Dendroctonus jeffreyi), Mountain pine beetle (D. ponderosae), and fir engraver (Scolytus ventralis) are a frequent disturbance in the study area that was modeled. Due to previous efforts, LANDIS-II has been calibrated to conditions in the Lake Tahoe Basin (Loudermilk et al. 2013, 2014, Scheller et al. 2019).
Using the CWHR classification scheme, we developed a rule set for LANDIS-II biomass outputs that would approximate CWHR habitats (Appendix 1). For this study, we modeled change in tree-dominated and shrub-dominated CWHR habitats (Fig. 1A). Areas that would be classified as aquatic, developed, or barren following CWHR classification rules were excluded from the landscape analysis because these areas were not projected to change over time. In addition, LANDIS-II does not model the herbaceous community, and we classified herbaceous-dominated habitat structure according to the woody vegetation component (i.e., tree or shrub-dominated) or as NULL following recent disturbances that had removed all woody vegetation. The shrub component of different habitat types can vary, and we classified an area as shrub dominated when ≥75% of the total biomass was attributed to shrubs (Appendix 1). This biomass threshold was chosen based on LANDIS-II biomass estimates of the current landscape compared to current classification of CWHR shrub-dominated habitats. Species composition in tree-dominated habitats followed the CWHR classification scheme but relied on species-specific biomass estimates instead of estimates of canopy cover. Species-specific biomass estimates produced by LANDIS-II to classify composition included: white fir (Abies concolor), red fir (A. magnifica), lodgepole pine (Pinus contorta), Jeffrey pine (P. jeffreyi), incense-cedar (Calocedrus decurrens), sugar pine (Pinus lambertiana), mountain hemlock (Tsuga mertensiana), western white pine (P. monticola), and whitebark pine (P. albicaulis). Aspen (Populus tremuloides) is the only hardwood species that occurs regularly within the study area and was the only tree-dominated habitat modeled that was not classified as conifer. Based on this rule set, we identified eight CWHR habitat types in our study area: White Fir (WFR), Red Fir (RFR), Jeffrey Pine (JPN), Lodgepole Pine (LPN), Sierra Mixed Conifer (SMC), Subalpine Conifer (SCN), Aspen (ASP), and Montane Chaparral (MCP).
For conifer dominated habitat types, we assigned each habitat type a seral stage based on CWHR size class descriptions (Fig. 1B). There are five CHWR size classes that influence habitat suitability for wildlife based on the mean diameter at breast height (DBH) of predominant trees. Seedling trees are the first size class and are defined by trees with a mean diameter < 2.5 cm, followed by sapling trees (size class 2: 2.5-15 cm), pole trees (size class 3: 15-28 cm), small trees (size class 4: 28-61 cm), and med/large trees (size class 5: > 61 cm). We estimated these size classes by combining estimates from LANDIS-II for the biomass of different species age cohorts with data from Forest Inventory and Analysis (FIA; Burrill et al. 2021) on the distribution of stem sizes (for equivalent species and age) in trees that have been measured in the northern Sierra Nevada ecoregion where Lake Tahoe is situated. We fit the Weibull distribution to FIA tree data for species-age groups that were equivalent to each LANDIS-II cohort. We assumed that the LANDIS-II cohorts would follow a similar size distribution to what was observed in the FIA data and distributed the LANDIS-II biomass estimates among two-inch DBH height bins (i.e., into a 0-5 cm bin, a 5-10 cm bin, etc.). To assign a pixel to a CHWR size class, we used the DBH of the 95th percentile tree in our estimated distribution and compared that to the CHWR size classes as described above (classes 1 through 5 being < 2.5 cm; 2.5-15 cm; 15-28 cm; 28-61 cm; and > 61 cm). We assessed different approaches, specifically, we also completed the classification using the size of the 90th percentile tree and the 99th percentile tree, and we found that using the 95th percentile yielded a CHWR in year 1 (2010) of the LANDIS-II model run that most closely reflected our baseline habitat distribution in the Lake Tahoe Basin. We considered this method reliable in producing three distinct tree-dominated seral stages: early (tree size approximating size classes 1-3), mid (tree size approximating size class 4), and late (tree size approximating size class 5).
Canopy cover in early seral habitat did not have a predictable relationship with canopy cover estimates, but for mid and late conifer-dominated habitats, we additionally assigned each pixel a canopy cover estimate (open, moderate, closed) based on CWHR canopy classes (Fig. 1C). Using the biomass bins described above, we again used the FIA data for average stem to biomass relationships at different size classes to translate biomass values into stem counts for each size bin (Evans et al. 2022). This process gave us an estimated size distribution of trees on each pixel as well as the estimated number of trees per pixel, which we translated into an estimate of trees per acre (TPA) to estimate canopy cover. We based our canopy thresholds on estimates of TPA whereby mid and late seral habitats with < 50 TPA were categorized as open, 51-500 TPA as moderate, and > 500 TPA as closed. Using these methods, we assigned each 1 ha pixel a CWHR habitat (composition, size class, canopy class). Although the CWHR classification scheme separates open from sparsely covered habitats, we combined these two into one category creating three canopy classes that are ecologically important and are used to establish guidelines for management. Descriptions of CWHR habitats modeled and the number of hectares of each habitat combination produced are presented in Appendix 2.
Simulated management scenarios
Management scenarios simulated in this study were selected by the Lake Tahoe Basin West Restoration Partnership to better understand the societal benefits of increasing the pace and scale of forest restoration (Table 1). Under the suppression-only (Suppress) scenario no forest management activities were modeled other than active fire suppression (i.e., if a fire started, it was suppressed). Under the wildland-urban interface (WUI) scenario, in addition to fire suppression, forest thinning (both mechanical and hand treatment) was modeled in the vegetated area adjoining structures and evacuation routes (which accounts for ~58% of the landscape). Similar to the WUI scenario, the thin scenario (Thin) modeled high levels of forest thinning in the WUI, as well as some treatments in the general forest and surrounding wilderness areas. The WUI and thin scenarios thus differed in the amount and extent of treatment. The two remaining scenarios were modeled with the use of fire as a management tool; both modeled moderate levels of thinning in the WUI while allowing for some prescribed burns and managed wildfire ignitions outside of the WUI. The fire scenario (Fire) however, simulated moderate levels of prescribed and managed fire, whereas the Fire+ scenario simulated high use of prescribed and managed fire. Information on the parameterization of the model is available here: https://github.com/LANDIS-II-Foundation/Project-Lake-Tahoe-2017
For each of the five scenarios, annual change in forest conditions were simulated from 2010-2110 using climate data from the CanESM2 global change model based on the relative concentration pathway 4.5. Each scenario run was simulated 10 times (i.e., 10 replicates) to quantify the mean and variation in change in habitat. At each 10-year timestep, we assigned CWHR habitat types to each spatial output (5 scenarios X 10 replicates X 10 decadal time steps) resulting in 500 spatially explicit habitat maps at 1-ha resolution.
Simulating wildlife-habitat outcomes
We extracted data from CWHR (https://www.wildlife.ca.gov/Data/CWHR) on wildlife-habitat relationships for each vertebrate species known to occur in the Lake Tahoe Basin (Schlesinger and Romsos 2000). We excluded any species from the analysis that relied primarily on aquatic habitats (e.g., amphibians and waterfowl). This resulted in a list of 135 birds, 51 mammals, and 5 reptiles. Of these 191 species, we were unable to simulate change in reproductive habitat for 23 birds and 2 mammals because they are not known to breed in the study area, or only breed in urban environments. Additionally, we excluded species in which their reproductive habitat was rare (< 5% of the landscape) including the Loggerhead Shrike (Lanius ludovicianus), Bullock’s Oriole (Icterus bullockii), White-crowned Sparrow (Zonotrichia leucophrys), Tree Swallow (Tachycineta bicolor), Lark Sparrow (Chondestes grammacus), and Gray-crowned Rosy Finch (Leucosticte tephrocotis). This resulted in 159 species in which a change in the value of habitat for reproductive purposes was modeled (Appendix 3).
For each modeled species and every habitat map (N = 500), we assigned a habitat-suitability ranking of high (1.00), moderate (0.66), low (0.33), or unsuitable (0.00) for reproduction based on the CWHR database for each 1-ha pixel (Fig. 2A). Because our translation from LANDIS II outputs (biomass by species by age cohort) to CWHR habitat resulted in the decision to combine certain CWHR habitats (i.e., sparse and open canopy classes were combined), we assigned the maximum value of reproductive habitat (hereafter habitat) for the combined categories (Appendix 3). From each map, we generated three metrics: (1) the total sum of habitat suitability values on the landscape for each species, (2) the proportion of the landscape classified as high-value habitat (i.e., suitability = 1; Fig. 2B), and (3) the average size of habitat patches of high reproductive value using ClassStat function in package SDMTools (McGarigal et al. 2002) in Program R version 3.6.1 (R Core Team 2018). For each species*scenario*replicate*timestep, we compared the computed metric to baseline starting conditions in 2010. For each metric (overall habitat quality, proportion of landscape considered high-quality habitat, average patch size of high-quality habitat) at each scenario*replicate*timestep, we summed the number of species in which each metric was predicted to increase and decrease. Because these metrics always fluctuate due to the dynamic nature of the model, we chose an arbitrary threshold of > 30% change to quantify more significant impacts on species and quantified these species separately.
RESULTS
Changes in habitat type and composition
At the start of our simulations, the modeled landscape was dominated by Jeffrey Pine (22%), White Fir (20%), Sierra Mixed Conifer (15%), and Montane Chaparral (21%) habitat types, with Aspen, Lodgepole Pine, Red Fir, and Subalpine Conifer Habitat types each comprising less than 10% of the landscape (Fig. 3). Under all scenarios, except the Fire+ scenario, the amounts of White Fir and Sierra Mixed Conifer Habitat types were predicted to increase over time, becoming the dominant habitat types, whereas all other habitat types tended to decrease. In the Fire+ scenario, increased use of fire was predicted to have a stabilizing effect on all habitat types over time, with Montane Chapparal quickly becoming the most common habitat type. Compared to the other scenarios, the Fire+ scenario maintained a larger percentage of Red Fir and Jeffrey Pine habitat types across the landscape.
Similar to the predicted outcomes for habitat type composition, management scenarios had little influence on the proportion of the landscape comprised of early, mid, and late seral stages over time (Fig. 3). Of the conifer dominated habitat types, 63% of the cells were classified initially as mid-seral, 26% as late seral, and 11% as early seral (the latter of which primarily occurred in White Fir and Red Fir habitat types). Under all management scenarios, the amount of early and mid-seral forest decreased whereas late seral forest increased. In the Fire+ scenario, conversion of more areas to Montane Chapparal resulted in a lower proportions of conifer forest, with late seral habitat conditions plateauing at ~22,500 ha and the smallest percentage of early seral conditions.
Compared to habitat type composition and seral stage, forest cover was more strongly influenced by management actions that increased the rate and extent of fuel reduction activities (Fig. 3). Compared to the management scenario simulating fire suppression (Suppress), fuel reduction treatments concentrated in the WUI (WUI) and with only moderate fire use (Fire) had a stabilizing effect on the proportions of the landscape in which forest cover was classified as moderate and closed. Increasing the pace and scale of fuel reduction treatments (Thin) was projected to reduce the amount of closed habitat conditions relative to the Suppress scenario but resulted in the largest proportion of the landscape being comprised of moderate forest cover. With the simulated increase in use of fire in the Fire+ scenario, forest cover across the landscape was projected to become sparser, with the most limited area of closed forest conditions.
Changes in suitability of reproductive habitat for wildlife
Of the 159 modeled species, 7 species were expected to experience a > 30% increase in the value of their habitat regardless of scenario: American Kestrel (Falco sparverius), Great Horned Owl (Bubo virginianus), Lewis’s Woodpecker (Melanerpes lewis), Williamson’s Sapsucker (Sphyrapicus thyroideus), Purple Martin (Progne subis), Western Bluebird (Sialia mexicana), and Evening Grosbeak (Coccothraustes vespertinus). Four species were predicted to lose over 30% of their reproductive habitat regardless of scenario including the Mountain Bluebird (Sialia currucoides), Western Meadowlark (Sturnella neglecta), House Finch (Carpodacus mexicanus), and mountain beaver (Aplodontia rufa).
For the remaining species the value of the landscape for reproductive purposes, both in the amount and configuration, differed by scenario. In terms of the overall value of reproductive habitat, all scenarios except the Fire+ scenario, led to outcomes that resulted in slightly more species experiencing increases in this metric than decreases (Fig. 4A, B). Under the Fire+ scenario, more species were predicted to experience declines in the overall amount of habitat for reproduction, although under this scenario the number of species experiencing significant change (i.e., > 30%) was minimized relative to the other scenarios (Fig. 4B).
In terms of high-quality reproductive habitat, all scenarios tended to have relatively neutral impacts on the proportion of their habitat categorized as high value. In the first half of the century, the Fire+ scenario resulted in increases in this metric for the largest number of species, whereas the Suppress scenario resulted in the least number of species (Fig. 4C). In the second half of the century, differences in this metric between scenarios were minimal and largely neutral. Similar to the results for the impact of scenarios on the overall habitat quantity of individual species, the Fire+ scenario tended to reduce the number of species experiencing more significant fluctuation in this metric during the first half of the century (Fig. 4D). For those species with > 30% fluctuation in the proportion of their high-quality reproductive habitat, more species were predicted to see increases in this metric relative to decreases, for all scenarios (Fig. 4D). Although the overall proportion of high-quality habitat was predicted to increase for as many species as it decreased, the average patch size of individual patches of this habitat was predicted to decrease for most of the species modeled, most notably under the Fire+ scenario (Fig. 4E, F). Differences between all other scenarios was minor and the variance in replicates overlapped, but the Thin scenario was predicted to lead to the most favorable biodiversity outcomes in the size of high-quality patches (Fig. 4C).
As patches of high-quality reproductive habitat were predicted to decrease over time regardless of scenario, we grouped modeled species based on their size and mobility and compared how each scenario was projected to impact the median size of high-quality reproductive patches for each grouping in 2040 (early century) and in 2070 the mid-century (Table 2, Appendix 3). For species with low mobility that tended to occupy small to moderate sized home ranges (≤ 40 ha; Manley and Schlesinger 2000), the median size of habitat patches decreased over the century regardless of scenario. These decreases were most dramatic under the Fire+ scenario both early and mid-century. Relative to the other scenarios, the Thin scenario was predicted to result in larger median patch sizes in the near term. In comparison, for larger-bodied species, the median patch size was predicted to be the smallest relative to other species groups and more similarly across time periods, with the Thin and Fire+ scenario leading to higher median patch sizes. For highly mobile species that typically occupy home ranges > 40 ha, patch size decreased over time regardless of scenario with more favorable outcomes again under the Thin and Fire+ scenario, both scenarios of which treated larger proportions of the landscape.
DISCUSSION
Recent extreme fire years in forested ecosystems across the globe have raised concerns regarding the potential impact of these climate-driven fires on biodiversity (Barlow et al. 2020, Kelly et al. 2020, Ward et al. 2020) and actions that can be taken to increase resilience of forests to future fires (Wintle et al. 2020, Stephens et al. 2021). In the seasonally dry forests of the western United States, forest restoration strategies that reduce tree densities and fuel loading have been shown to alter fire behavior and reduce fire severity (Stephens and Moghaddas 2005, Tubbesing et al. 2019). Our model results suggest that management regimes that incorporated more extensive use of fuel reduction treatments and fire can alter forest trajectories and habitat for biodiversity.
With the exception of a management regime incorporating more prescribed and managed wildfire (Fire+), our models indicated that forests surrounding Lake Tahoe will experience an increase in White Fir and Sierra Mixed Conifer and a decrease in stands dominated by Jeffrey pine and montane chapparal. This prediction is supported by previous research indicating that Jeffrey pine is adapted to a frequent-fire disturbance regime whereby frequent removal of small diameter trees and fire-intolerant tree species by low-intensity fire is needed to maintain this species on the landscape (Fulé et al. 2012). Our modeling results suggest that higher use of fire is needed to maintain Jeffrey Pine habitat in our study area. Previous research has also shown that red fir seedling establishment and growth are strongly related to periodic disturbance (Chappell and Agee 1996) and this may explain why the Fire+ scenario was also the only scenario predicted to maintain Red Fir habitat. Climate change is predicted to have a negative impact on red fir (Meng et al. 2015) as establishment of red fir seedlings in influenced by soil moisture and post-fire temperatures and maintaining Red Fir habitat will be a future management challenge. Although maintaining a more even mix of habitat types, increased use of fire under the Fire+ scenario led to the highest proportion of montane chapparal. Many non-conifer vegetation communities have been negatively impacted by fire suppression including montane chapparal and riparian communities, including aspen (White and Long 2019), and our modeling results indicate that higher use of fire may be one way to ensure persistence of these habitat types in the Lake Tahoe Basin.
Whereas frequent use of fire appeared to be important in maintaining a diversity of vegetative communities, the area being treated led to larger differences among scenarios in the density of forest canopy. Increasing levels of disturbance resulted in landscapes with a higher percentage of the landscape comprised of moderate tree densities and associated canopy (Thin scenario) or low tree densities and open canopy (Fire+ scenario). Over the century, present day areas with open canopy were predicted to transition to areas with moderate canopy cover consistent with forest growth and succession with limited disturbance and competition for resources. This trajectory is similar to that observed over that last century with increasing stand densities of smaller-sized trees and less structural variability (Taylor 2004). Landscape restoration strategies incorporating more extensive fuel reduction resulted in forests with more moderate tree densities that are less prone to stand-replacing fires (Agee and Skinner 2005) and more resilient to a changing climate (North et al. 2019), but that also may be less conducive to species preferring closed canopy conditions.
One of our main objectives was to understand the impact of different landscape management scenarios on wildlife in the Lake Tahoe Basin based on wildlife-habitat associations. Our modeling results suggest that over the next century changes in wildlife habitat will occur in quantifiable ways, but predictions for most species indicate that they will experience habitat fluctuations that are less than 30% of their current habitat regardless of management approach, suggesting that over the long-term the trajectory of forest change will have an overall neutral effect on biodiversity. Although the Fire+ scenario was predicted to promote greater representation of different habitat types, it was predicted to result in a reduction in the overall value of the habitat for many species despite maintaining more consistent amounts of high-quality reproductive habitat than the other scenarios (Fig. 4). This suggests that in our study area, structural variables of forest habitat were more influential in maintaining habitat for biodiversity than forest composition.
The reduced frequency and extent of fire have caused an increase in forest density, a shift to more fire-sensitive species and large-scale homogenization of forest conditions (Taylor 2001, Safford and Stevens 2017). Over the next century, our simulations project that disturbances altering forest structure will break up the landscape regardless of scenario, reducing the current size of large contiguous forest patches. The distribution of habitat across landscape will impact species differently depending on the distribution of needed resources and their ability to transverse landscapes (Kindlmann and Burel 2008). Based on the size of high-valued reproductive habitat patches, we attempted to predict how different management approaches were likely to impact different groupings of wildlife based on their typical home-range size and mobility (Manley and Schlesinger 2000). We acknowledge that the habitat matrix (i.e., inter-patch habitat) can be as important as the size of habitat patches (Tischendorf and Fahrig 2000), but this is beyond the scope of this study. In general, we found that patches of contiguous high-quality reproductive habitat would decrease for the majority of species, most notably under the Fire+ scenario (Fig. 4D, E). Management scenarios that were simulated at greater extents (i.e., the Thin and Fire+ scenarios) tended to result in larger patch sizes for the more mobile species and those with larger home ranges. This result in combination with the metric of overall habitat quality for wildlife indicates that management strategies are likely to have idiosyncratic effects of wildlife species, but biodiversity in this disturbance-prone system is fairly resilient to change (see also Sollmann et al. 2015, Kelt et al. 2017). However, in our model we assume that habitat suitability is strongly linked to forest composition and structure, and we do not account for other abiotic and biotic factors that determine the capacity of a given habitat to support the needs of a species.
CONCLUSIONS
Change is predicted, and management can have an impact on the trajectory and speed of change for habitat types and associated species. Choice of best management approach is a societal decision (Abelson et al. 2022) and will depend on prioritization of resources and outcomes, of which biodiversity is just one. Our results are consistent with other studies (Fontaine and Kennedy 2012, Stephens et al. 2014, Stevens et al. 2016) suggesting that different management actions will alter habitat suitability for biodiversity, but that the effects will be species-dependent with compensatory effects on species richness. However, our simulations are based on a representative concentration pathway (rcp 4.5) that does not match our current global emissions (Schwalm et al. 2020). It is likely that use of a higher greenhouse gas emissions scenario would increase the amount of fire on the landscape, leading to more frequent disturbance and landscape conditions most similar to the Fire+ scenario. We have only considered one metric for measuring biodiversity without considering the functional role of individual species (Caro 2010, Slauson et al. 2022), which may be as important in sustaining ecosystem function as maintaining current species richness. Ensuring the sustainability of forested ecosystems into the future will continue to be a challenge for managers and society, and integrated frameworks are needed to assess the value of future landscapes (Stevens et al. 2016, Abelson et al. 2022).
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.AUTHOR CONTRIBUTIONS
RS, AK, and CM formulated the LANDIS model and computed the outputs necessary to implement the wildlife habitat model. AW, TH, AK, and AB developed the framework for habitat compilations. AW analyzed the data and wrote the manuscript with input from all authors. AW conceived the study and was in charge of overall direction and planning.
ACKNOWLEDGMENTS
We acknowledge the indigenous Native American people, the Washoe, who originally lived around the Lake Tahoe Basin and pay our respect to them, their culture, and their Elders, past, present, and future. Our research was informed by continual discussions with many dedicated people who actively participated the in the Lake Tahoe West Restoration Partnership, and it is through these conversations that progress can be best made. In particular, we acknowledge the contributions of P. Manley, J. Long, S. Di Vittori to project management, vision, and oversight and to members of the wildlife team including: S. Coppeto, W. Brennan, K. Slauson, S. Britting who all shared their individual perspectives and experiences. We thank M. K. Faber for producing highly effective figures that speak 1000 words and to 2 anonymous reviewers who greatly improved this manuscript.
DATA AVAILABILITY
Parameters used in the LANDIS modeling are available here: https://github.com/LANDIS-II-Foundation/Project-Lake-Tahoe-2017, which has been archived here: https://doi.org/10.5281/zenodo.4644579
LITERATURE CITED
Abelson, E., K. Reynolds, A. M. White, J. W. Long, C. Maxwell, and P. N. Manley. 2022. Evaluating pathways to social and ecological resilience. Ecology and Society, in press.
Agee, J. K., and C. N. Skinner. 2005. Basic principles of forest fuel reduction treatments. Forest Ecology and Management 211:83-96. https://doi.org/10.1016/j.foreco.2005.01.034
Barbour, M., E. Kelley, P. Maloney, D. Rizzo, E. Royce, and J. Fites-Kaufmann. 2002. Present and past old-growth forests of the Lake Tahoe Basin, Sierra Nevada, US. Journal of Vegetation Science 13:461-472. https://doi.org/10.1111/j.1654-1103.2002.tb02073.x
Barlow, J., E. Berenguer, R. Carmenta, and F. França. 2020. Clarifying Amazonia’s burning crisis. Global Change Biology 26:319-321. https://doi.org/10.1111/gcb.14872
Beaty, R. M., and A. H. Taylor. 2007. Fire disturbance and forest structure in old-growth mixed conifer forests in the northern Sierra Nevada, California. Journal of Vegetation Science 18:879-890. https://doi.org/10.1111/j.1654-1103.2007.tb02604.x
Burrill, E. A., A. M. DiTommaso, J. A. Turner, S. A. Pugh, J. Menlove, G. Christiansen, C. J. Perry, and B. L. Conkling. 2021. The forest inventory and analysis database: database description and user guide version 9.0 for Phase 2. U.S. Department of Agriculture, Forest Service, Washington, D.C., USA. http://www.fia.fs.fed.us/library/database-documentation/
Caro, T. 2010. Conservation by proxy: indicator, umbrella, keystone, flagship, and other surrogate species. Island, Washington, D.C., USA.
Chappell, C. B., and J. K. Agee. 1996. Fire severity and tree seedling establishment in Abies magnifica forests, southern Cascades, Oregon. Ecological Applications 6:628-640. https://doi.org/10.2307/2269397
Evans, S. G., T. G. Holland, J. W. Long, C. Maxwell, R. M. Scheller, E. Patrick, and M. D. Potts. 2022. Modeling the risk reduction benefit of forest management using a case study in the Lake Tahoe Basin. Ecology and Society 27(2):18. https://doi.org/10.5751/ES-13169-270218
Fontaine, J. B., and P. L. Kennedy. 2012. Meta-analysis of avian and small-mammal response to fire severity and fire surrogate treatments in US fire-prone forests. Ecological Applications 22:1547-1561. https://doi.org/10.1890/12-0009.1
Fulé, P. Z., J. E. Crouse, J. P. Roccaforte, and E. L. Kalies. 2012. Do thinning and/or burning treatments in western USA ponderosa or Jeffrey pine-dominated forests help restore natural fire behavior? Forest Ecology and Management 269:68-81. https://doi.org/10.1016/j.foreco.2011.12.025
Kelly, L. T., K. M. Giljohann, A. Duane, N. Aquilué, S. Archibald, E. Batllori, A. F. Bennett, S. T. Buckland, Q. Canelles, M. F. Clarke, M.-J. Fortin, V. Hermoso, S. Herrando, R. E. Keane, F. K. Lake, M. A. McCarthy, A. Morán-Ordóñez, C. L. Parr, J. G. Pausas, T. D. Penman, A. Regos, L. Rumpff, J. L. Santos, A. L. Smith, A. D. Syphard, M. W. Tingley, and L. Brotons. 2020. Fire and biodiversity in the Anthropocene. Science 370(6519). https://doi.org/10.1126/science.abb0355
Kelt, D. A., R. Sollmann, A. M. White, S. L. Roberts, and D. H. Van Vuren. 2017. Diversity of small mammals in the Sierra Nevada: filtering by natural selection or by anthropogenic activities? Journal of Mammalogy 98:85-93. https://doi.org/10.1093/jmammal/gyw158
Kindlmann, P., and F. Burel. 2008. Connectivity measures: a review. Landscape Ecology 23:879-890. https://doi.org/10.1007/s10980-008-9245-4
Knapp, E. E., C. N. Skinner, M. P. North, and B. L. Estes. 2013. Long-term overstory and understory change following logging and fire exclusion in a Sierra Nevada mixed-conifer forest. Forest Ecology and Management 310:903-914. https://doi.org/10.1016/j.foreco.2013.09.041
Larson, A. J., and D. Churchill. 2012. Tree spatial patterns in fire-frequent forests of western North America, including mechanisms of pattern formation and implications for designing fuel reduction and restoration treatments. Forest Ecology and Management 267:74-92. https://doi.org/10.1016/j.foreco.2011.11.038
Loudermilk, E. L., R. M. Scheller, P. J. Weisberg, J. Yang, T. E. Dilts, S. L. Karam, and C. Skinner. 2013. Carbon dynamics in the future forest: the importance of long-term successional legacy and climate-fire interactions. Global Change Biology 19:3502-3515. https://doi.org/10.1111/gcb.12310
Loudermilk, E. L., A. Stanton, R. M. Scheller, T. E. Dilts, P. J. Weisberg, C. Skinner, and J. Yang. 2014. Effectiveness of fuel treatments for mitigating wildfire risk and sequestering forest carbon: a case study in the Lake Tahoe Basin. Forest Ecology and Management 323:114-125. https://doi.org/10.1016/j.foreco.2014.03.011
Lutz, J. A., A. J. Larson, and M. E. Swanson. 2018. Advancing fire science with large forest plots and a long-term multidisciplinary approach. Fire 1:5. https://doi.org/10.3390/fire1010005
Lydersen, J. M., M. P. North, and B. M. Collins. 2014. Severity of an uncharacteristically large wildfire, the Rim Fire, in forests with relatively restored frequent fire regimes. Forest Ecology and Management 328:326-334. https://doi.org/10.1016/j.foreco.2014.06.005
Manley, P. N., and M. D. Schlesinger. 2000. Designation of focal vertebrate species for the Lake Tahoe Basin. Pages L1-L6 in D. D. Murphy and C. M. Knopp, editors. Lake Tahoe watershed assessment. Volume II. General Technical Report PSW-GTR-176. US Department of Agriculture Forest Service, Albany, California, USA. https://www.fs.fed.us/psw/publications/documents/psw_gtr176/psw_gtr176_appendixL.pdf
Mayer, K. E., and W. F. Laudenslayer. 1988. A guide to wildlife habitats of California. California Department of Forestry and Fire Protection, Sacramento, California, USA.
McGarigal, K., S. A. Cushman, M. C. Neel, and E. Ene. 2002. FRAGSTATS: spatial pattern analysis program for categorical maps. University of Massachusetts, Amherst, Massachusetts, USA.
Meng, R., P. E. Dennison, C. Huang, M. A. Moritz, and C. D’Antonio. 2015. Effects of fire severity and post-fire climate on short-term vegetation recovery of mixed-conifer and red fir forests in the Sierra Nevada Mountains of California. Remote Sensing of Environment 171:311-325. https://doi.org/10.1016/j.rse.2015.10.024
Mladenoff, D. J. 2004. LANDIS and forest landscape models. Ecological Modelling 180:7-19. https://doi.org/10.1016/j.ecolmodel.2004.03.016
North, M., A. Brough, J. Long, B. Collins, P. Bowden, D. Yasuda, J. Miller, and N. Sugihara. 2015. Constraints on mechanized treatment significantly limit mechanical fuels reduction extent in the Sierra Nevada. Journal of Forestry 113:40-48. https://doi.org/10.5849/jof.14-058
North, M., B. M. Collins, and S. Stephens. 2012. Using fire to increase the scale, benefits, and future maintenance of fuels treatments. Journal of Forestry 110:392-401. https://doi.org/10.5849/jof.12-021
North, M., and P. N. Manley. 2012. Managing Sierra Nevada forests. US Department of Agriculture, Forest Service, Pacific Southwest Research Station, Albany, California, USA. https://doi.org/10.2737/PSW-GTR-237
North, M., P. Stine, K. O’Hara, W. Zielinski, and S. Stephens. 2009. An ecosystem management strategy for Sierran mixed-conifer forests. General Technical Report PSW-GTR-220 (Second printing, with addendum). US Department of Agriculture, Forest Service, Pacific Southwest Research Station, Albany, California, USA. https://doi.org/10.2737/PSW-GTR-220
North, M. P., J. T. Stevens, D. F. Greene, M. Coppoletta, E. E. Knapp, A. M. Latimer, C. M. Restaino, R. E. Tompkins, K. R. Welch, R. A. York, D. J. N. Young, J. N. Axelson, T. N. Buckley, B. L. Estes, R. N. Hager, J. W. Long, M. D. Meyer, S. M. Ostoja, H. D. Safford, K. L. Shive, C. L. Tubbesing, H. Vice, D. Walsh, C. M.Werner, and P. Wyrsch. 2019. Tamm review: reforestation for resilience in dry western U.S. forests. Forest Ecology and Management 432:209-224. https://doi.org/10.1016/j.foreco.2018.09.007
R Core Team. 2018. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/.
Safford, H. D., and J. T. Stevens. 2017. Natural range of variation for yellow pine and mixed-conifer forests in the Sierra Nevada, southern Cascades, and Modoc and Inyo National Forests, California, USA. General Technical Report PSW-GTR-256. US Department of Agriculture, Forest Service, Pacific Southwest Research Station, Albany, California, USA. https://doi.org/10.2737/PSW-GTR-256
Scheller, R. M., J. B. Domingo, B. R. Sturtevant, J. S. Williams, A. Rudy, E. J. Gustafson, and D. J. Mladenoff. 2007. Design, development, and application of LANDIS-II, a spatial landscape simulation model with flexible temporal and spatial resolution. Ecological Modelling 201:409-419. https://doi.org/10.1016/j.ecolmodel.2006.10.009
Scheller, R., A. Kretchun, T. J. Hawbaker, and P. D. Henne. 2019. A landscape model of variable social-ecological fire regimes. Ecological Modelling 401:85-93. https://doi.org/10.1016/j.ecolmodel.2019.03.022
Schlesinger, M. D., and J. S. Romsos. 2000. Vertebrate species of the Lake Tahoe Basin. Pages G1-G14 in D. D. Murphy and C. M. Knopp, editors. Lake Tahoe watershed assessment. Volume II. General Technical Report PSW-GTR-176. US Department of Agriculture Forest Service, Albany, California, USA. https://www.fs.fed.us/psw/publications/documents/psw_gtr176/psw_gtr176_appendixG.pdf
Schwalm, C. R., D. N. Huntzinger, A. M. Michalak, K. Schaefer, J. B. Fisher, Y. Fang, and Y. Wei. 2020. Modeling suggests fossil fuel emissions have been driving increased land carbon uptake since the turn of the 20th Century. Scientific Reports 10(1):9059. https://doi.org/10.1038/s41598-020-66103-9
Slauson, K., B. Howard, A. M. White, C Maxwell, and T. Holland. 2022. Evaluating the effects of alternative landscape management scenarios on three old forest associated predators over 100-years in the fire prone forests of the Sierra Nevada, USA. Ecology and Society, in press.
Smucker, K. M., R. L. Hutto, and B. M. Steele. 2005. Changes in bird abundance after wildfire: importance of fire severity and time since fire. Ecological Applications 15:1535-1549. https://doi.org/10.1890/04-1353
Sollmann, R., A. M. White, B. Gardner, and P. N. Manley. 2015. Investigating the effects of forest structure on the small mammal community in frequent-fire coniferous forests using capture-recapture models for stratified populations. Mammalian Biology 80:247-254. https://doi.org/10.1016/j.mambio.2015.03.002
Stephens, S. L., J. K. Agee, P. Z. Fulé, M. P. North, W. H. Romme, T. W. Swetnam, and M. G. Turner. 2013. Managing forests and fire in changing climates. Science 342:41-42. https://doi.org/10.1126/science.1240294
Stephens, S. L., S. W. Bigelow, R. D. Burnett, B. M. Collins, C. V. Gallagher, J. Keane, D. A. Kelt, M. P. North, L. J. Roberts, P. A. Stine, and D. H. Van Vuren. 2014. California Spotted Owl, songbird, and small mammal responses to landscape fuel treatments. BioScience 64:893-906. https://doi.org/10.1093/biosci/biu137
Stephens, S. L., A. LeRoy Westerling, M. D. Hurteau, M. Z. Peery, C. A. Schultz, and S. Thompson. 2021. Undesirable outcomes in seasonally dry forests. Frontiers in Ecology and the Environment 19:87-88. https://doi.org/10.1002/fee.2309
Stephens, S. L., and J. J. Moghaddas. 2005. Experimental fuel treatment impacts on forest structure, potential fire behavior, and predicted tree mortality in a California mixed conifer forest. Forest Ecology and Management 215:21-36. https://doi.org/10.1016/j.foreco.2005.03.070
Stevens, J. T., B. M. Collins, J. W. Long, M. P. North, S. J. Prichard, L. W. Tarnay, and A. M. White. 2016. Evaluating potential trade-offs among fuel treatment strategies in mixed-conifer forests of the Sierra Nevada. Ecosphere 7:e01445. https://doi.org/10.1002/ecs2.1445
Swanson, M. E., J. F. Franklin, R. L. Beschta, C. M. Crisafulli, D. A. DellaSala, R. L. Hutto, D. B. Lindenmayer, and F. J. Swanson. 2011. The forgotten stage of forest succession: early-successional ecosystems on forest sites. Frontiers in Ecology and the Environment 9(2):117-125. https://doi.org/10.1890/090157
Taylor, A. H. 2001. Fire regimes and forest changes in mid and upper montane forests of the southern Cascades, Lassen Volcanic National Park, California, U.S.A. Journal of Biogeography 27:87-104. https://doi.org/10.1046/j.1365-2699.2000.00353.x
Taylor, A. H. 2004. Identifying forest reference conditions on early cut-over lands, Lake Tahoe Basin, USA. Ecological Applications 14:1903-1920. https://doi.org/10.1890/02-5257
Tingley, M. W., V. Ruiz-Gutiérrez, R. L. Wilkerson, C. A. Howell, and R. B. Siegel. 2016. Pyrodiversity promotes avian diversity over the decade following forest fire. Proceedings of the Royal Society B: Biological Sciences 283:20161703. https://doi.org/10.1098/rspb.2016.1703
Tischendorf, L., and L. Fahrig. 2000. On the usage and measurement of landscape connectivity. Oikos 90:7-19. https://doi.org/10.1034/j.1600-0706.2000.900102.x
Tubbesing, C. L., D. L. Fry, G. B. Roller, B. M. Collins, V. A. Fedorova, S. L. Stephens, and J. J. Battles. 2019. Strategically placed landscape fuel treatments decrease fire severity and promote recovery in the northern Sierra Nevada. Forest Ecology and Management 436:45-55. https://doi.org/10.1016/j.foreco.2019.01.010
Ward, M., A. I. Tulloch, J. Q. Radford, B. A. Williams, A. E. Reside, S. L. Macdonald, H. J. Mayfield, M. Maron, H. P. Possingham, S. J. Vine, J. L. O’Connor, E. J. Massingham, A. C. Greenville, J. C. Z. Woinarski, S. T. Garnett, M. Lintermans, B. C. Scheele, J. Carwardine, D. G. Nimmo, D. B. Lindenmayer, R. M. Kooyman, J. S. Simmonds, L. J. Sonter, and J. E. M. Watson. 2020. Impact of 2019-2020 mega-fires on Australian fauna habitat. Nature Ecology and Evolution 4:1321-1326. https://doi.org/10.1038/s41559-020-1251-1
White, A. M., and J. W. Long. 2019. Understanding ecological contexts for active reforestation following wildfires. New Forests 50:41-56. https://doi.org/10.1007/s11056-018-9675-z
White, A. M., P. N. Manley, G. L. Tarbill, T. W. Richardson, R. E. Russell, H. D. Safford, and S. Z. Dobrowski. 2016. Avian community responses to post-fire forest structure: implications for fire management in mixed conifer forests. Animal Conservation 19:256-264. https://doi.org/10.1111/acv.12237
White, A. M., E. F. Zipkin, P. N. Manley, and M. D. Schlesinger. 2013. Simulating avian species and foraging group responses to fuel reduction treatments in coniferous forests. Forest Ecology and Management 304:261-274. https://doi.org/10.1016/j.foreco.2013.04.039
Wintle, B. A., S. Legge, and J. C. Woinarski. 2020. After the megafires: what next for Australian wildlife? Trends in Ecology and Evolution 35:753-757. https://doi.org/10.1016/j.tree.2020.06.009
Young, D. J., J. T. Stevens, J. M. Earles, J. Moore, A. Ellis, A. L. Jirka, and A. M. Latimer. 2017. Long-term climate and competition explain forest mortality patterns under extreme drought. Ecology Letters 20:78-86. https://doi.org/10.1111/ele.12711
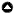
Fig. 1

Fig. 1. Location of the Lake Tahoe Basin depicting (A) the distribution of modeled habitat types, (B) seral stage, and (C) canopy cover class at the start of LANDIS-II modeling simulations in 2010.
Fig. 2

Fig. 2. An example of the outputs used for assessing wildlife metrics: (A) total value of reproductive habitat on the landscape, and (B) patches of high-quality reproductive habitat. This figure is of replicate 1 of the Northern flying squirrel (Glaucomys sabrinus) under the Thin scenario in 2070.
Fig. 3

Fig. 3. Changes in forest type, seral stage, and cover class over the period of modeling under each management scenario. Note forest types: White Fir (WFR), Red Fir (RFR), Jeffrey Pine (JPN), Lodgepole Pine (LPN), Sierra Mixed Conifer (SMC), Subalpine Conifer (SCN), Aspen (ASP), and Montane Chaparral (MCP).
Fig. 4

Fig. 4. The number of species projected to experience positive and negative changes in (A) the value of reproductive habitat, (B) the number of species in which change in A is > 30%, (C) proportion of the landscape considered high quality, (D), the number of species in which change in C is > 30%, (E) the average size of patches of high-quality habitat, and (F) the number of species in which change in E is > 30% under each management scenario.
Table 1
Table 1. Names and descriptions of the five management scenarios modeled.
| Scenario name | Modeled management actions | Scenario intent |
| Suppress | Management actions were limited to suppressing all fire activity. | Understand the societal benefits of fire suppression. |
| Wildland-urban interface (WUI) | In addition to fire suppression, approximately 1.8% of the vegetated area was treated annually in the WUI and defense zone. | Understand the societal benefits of current management actions focused on reducing fuels around human-built structures and evacuation zones. |
| Thin | In addition to fire suppression and WUI-focused fuel reduction treatments, this scenario treated approximately 6.7% of the vegetated area annually including the forest beyond the WUI. | Understand the societal benefits of expanding the use of mechanical and hand-thinning into the general forest. |
| Fire | Management actions were focused on reducing fuels via prescribed fire and managed wildfire with approximately 4% of the vegetated area treated each year. | Understand the societal benefits of using fire-driven fuel reduction treatments in addition to mechanical and hand-thinning mechanisms. |
| Fire+ | Management actions were focused on reducing fuels via prescribed fire and managed wildfire with approximately 7.2% of the vegetated area treated each year. | Increase the amount of vegetated area treated each year via fire-driven mechanisms (75% of all fuel reductions) to better differentiate between the societal benefits of fuel reduction mechanisms and the area treated. |
Table 2
Table 2. Comparisons of the median and range of patch sizes in 2040 and 2070 for different ecological groups of taxa under the five management scenarios relative to baseline conditions.
| Group | Baseline (ha) | Scenario | 2040 (ha) | 2070 (ha) |
| Small-bodied | 10.8 (6.26-17.1) |
Suppress | 8.9 (2.9-10.9) | 7.8 (2.1-9.8) |
| WUI | 8.6 (3-10.7) | 7 (2.2-9.9) | ||
| Thin | 9.2 (3.2-11.7) | 7.1 (2.5-12.2) | ||
| Fire | 8.5 (3-10.5) | 7.2 (2.2-10.3) | ||
| Fire+ | 6.9 (2.4-11.2) | 6.8 (2.5-8.8) | ||
| Medium-bodied | 9.23 (6.42-17.2) |
Suppress | 5.1 (4.8-11) | 5.3 (3.1-9.7) |
| WUI | 5.7 (4.8-11.5) | 5.8 (3.2-13.1) | ||
| Thin | 6.8 (5-13.1) | 6.9 (3.7-13.7) | ||
| Fire | 5.4 (4.7-11.2) | 6 (3.1-13.4) | ||
| Fire+ | 6.9 (3.1-12) | 6.4 (2.8-9.4) | ||
| High mobility | 17.3 (8.96-36.8) |
Suppress | 10.8 (4.9-44.6) | 8 (5.8-76.4) |
| WUI | 11.2 (6.2-38.6) | 7.5 (6.6-65) | ||
| Thin | 12.9 (6.9-36.8) | 8.4 (7.1-53.7) | ||
| Fire | 10.9 (5.9-38.4) | 7.8 (6.3-65.2) | ||
| Fire+ | 12.5 (6.4-28.5) | 9.4 (5.2-25.1) | ||