






The following is the established format for referencing this article:
Schrenk, H., B. Magnússon, B. D. Sigurdsson, and W. zu Castell. 2022. Systemic analysis of a developing plant community on the island of Surtsey. Ecology and Society 27(1):35.ABSTRACT
Based on our recently developed method to quantify Gunderson and Holling's adaptive cycle, we provide a holistic analysis of a vascular plant community on the volcanic island of Surtsey between 2000 and 2018. We identify one complete adaptive cycle during the study period, which reflects the system's transition from a classic pioneer to a grassland community. Our results support the hypothesis that nutrients brought to the island by breeding gulls are the main driver of this development. The study period includes the beginning of a second cycle, which deviates from the pattern described in the metaphor. Indeed, the cycle's exploitation phase is interrupted by a simultaneous decline of all three systemic variables. We can trace this phenomenon back to a severe drought in 2012. Furthermore, the method allows us to establish the systemic role of individual species during the maturation process.INTRODUCTION
Analyzing the dynamics of change in ecosystems remains a challenging endeavor. Ecosystem characteristics are formed by physical and chemical processes, as well as the interactions of organisms inhabiting the system, their adaptation to the given environment, and the shaping of the latter.
Dynamics of ecosystem development have been extensively studied and several frameworks for qualitative (Bossel 1992, Holling 2001) and quantitative descriptions (Odum 1983, Ulanowicz 1986) have been proposed. One of the qualitative descriptions is given by Gunderson and Holling’s adaptive cycle metaphor (Gunderson and Holling 2002), centerpiece of their panarchy theory. According to the metaphor, system development is shaped by three comprehensive systemic properties: (1) the system’s potential available for future change, (2) the connectedness among its internal variables and processes, and (3) its resilience in the light of unpredicted perturbations. The interplay of these three properties defines four phases of varying degrees of predictability. Encountering a space of largely unexplored resources, a system will start to grow by making use of the available resources. During this period, ecosystems are dominated by so-called r-strategists, which are characterized by a high dispersal ability and low demands on their habitat (Pianka 1970). Following this terminology, this period is called “r-phase” or phase of exploitation. Typically, resilience is high, whereas potential and connectedness still have to be build-up. Over time, the system increases its inner level of organization and thereby its access to resources, for example, being reflected by the opening of new niches (Odling-Smee et al. 1996). However, the increased level of specialization results in growing rigidity, leading to a decrease in resilience. Following ecological terminology, this phase is called “K-phase” or phase of conservation. It is characterized by a high degree of activity and efficiency, resulting in high levels of potential and connectedness. Together, the r- and K-phases comprise a period of consolidation and predictability. At some point, a trigger will lead to the destruction of established structure during the so-called “Ω-phase.” Resources are being release and former connections are broken. Potential and connectedness strongly decline, while resilience increases. Eventually, processes of mobilization initiate the system’s α-phase or phase of reorganization. During this period, new opportunities are exploited, new components might enter the system, whereas others might get lost. The latter two phases are thus characterized by a high degree of stochasticity, admitting chance for creative change. With a set of components slowly settling in, the system starts into another r-phase. According to the metaphor, this alternation between growth and renewal, consolidation and creativity, allows a complex system to repeatedly adapt to a changing environment.
Due to its high degree of generality, the metaphor serves as a valuable framework for the development of a broad range of complex systems. However, it has its limits as well. Systems living passively with external variability and systems anticipating and manipulating variability may, for example, permanently remain in single phases (Gunderson and Holling 2002). Besides, the interplay of internal and external factors may generally result in the skipping or stretching of single phases. Although being intuitively plausible, the adaptive cycle has so far to a great extent eluded itself from being empirically validated because of the intrinsic difficulty in quantifying the underlying driving variable’s potential, connectedness, and resilience (see e.g., Burkhard et al. 2011, Fath et al. 2015). In Castell and Schrenk 2020, we presented a method to quantify the adaptive cycle. Taking advantage of the abstract framework of information theory, the method is universally applicable and requires only the most basic information about a system, namely the time series of its components’ abundance data. This opens a wide range of possibilities for application because to minimize disturbance of the system of observation, species abundance counts remain the most commonly found approach for ecosystem assessment.
Our aim is to provide a holistic analysis of the development of a vascular plant community on the island of Surtsey following the approach introduced in Castell and Schrenk 2020. In 1963, volcanic eruption formed an island southwest of Heimaey, one of Iceland’s offshore islands. The island being newly formed has been named Surtsey, and its geological and biological development have been closely monitored since then (Fridriksson 2005). As soon as two years after its formation, the first higher plant was observed on the island although it was not yet able to persist (Fridriksson 2005). By 1985, a stable community of pioneer plants had developed on the southern part of the volcanic island, where the terrain was sheltered from heavy winter seas and gulls had started to breed, bringing in further seeds and nutrients (Magnusson et al. 2014). Since then, communities of vascular plants have evolved through several stages, thus providing a valuable dataset to assess ecosystem development.
Analyzing the community of vascular plants on a permanent monitoring plot between 2000 and 2018, patterns of ecological succession can be identified, reflecting the transition of the community from early pioneers toward a stable grassland ecosystem. The system runs through one complete adaptive cycle and starts into another one during the study period. Our analysis supports the identification of breeding gulls as external drivers of the first system breakdown. The second cycle shows that significant changes of environmental conditions lead to deviations from the metaphor. Indeed, during the second cycle, the system’s exploitation phase is suddenly interrupted by a simultaneous decline of all three systemic variables. This phenomenon can be traced back to a severe drought.
METHODS
Computing the adaptive cycle
Our method of operationalizing the adaptive cycle has been presented in detail in Castell and Schrenk 2020. Hence, we confine ourselves to a brief review of the underlying idea at this point. A technical description including the explicit mathematical formulas can be found in Appendix 1.
Consider a system of interacting agents. Assume that the state of each agent is known for a fixed set of time points. We will call such data “abundance data,” although we do not restrict ourselves to abundance in the strict sense of the word. The only demands on the data are that they at least implicitly capture the qualitative nature of interaction patterns among the agents. In the case of an ecosystem, such data could, for example, be biomass or the number of individuals per species. In the case of an economic system, capital could quantify the abundance of a company. We now assume that every effective interaction leads to a transfer of information between the interacting agents and reveals itself in interdependencies between the corresponding time series of abundance data. Building on these assumptions, we use Schreiber’s transfer entropy (Schreiber 2000) to quantify interdependencies. Transfer entropy from agent Y to agent X measures the amount of randomness in the outcome of agent X’s abundance being explained by the knowledge of agent Y’s abundance. In other words: how much can we learn about the future of X knowing not only the past of X but the past of Y as well? We estimate transfer entropy between every ordered pair of agents. By considering the agents as nodes and information transfers as edges, the results can be represented in the form of a weighted, directed graph. Instead of the whole time series, we base network estimation on a certain time window only. By shifting this time window, we gain a series of networks, reflecting the temporal development of the system’s information structure. For simplicity, we refer to these networks as the system’s “information networks.” The total amount of information being transferred from a node will be denoted by its information “outflow” and by the amount of information being transferred to a node, i.e., its information “inflow.”
We defined potential, connectedness, and resilience as properties of the system’s information network. Hence, the series of networks gained in the first step of the method serves as a basis to determine the development of the three systemic variables, independently of the concrete instantiation of the system. Our definitions of potential and connectedness are inspired by Ulanowicz’s notions of capacity and ascendency (Ulanowicz et al. 2009). Potential is a measure of the average entropy in the information network (compare Cover and Thomas 2006). It thereby captures the potential available for change or, in other words, its range of future options possible. It is high in the case of many equally weighted edges in the information network. Intuitively, a diverse interaction structure, being reflected in a broad range of information transfers, allows the system to shift its focus with respect to environmental variation.
Connectedness measures the average mutual information between the information outflow of one node and the information inflow of another node, being realized by edges in the network (compare Cover et al. 2006). As such, it quantifies the mutual constraints and dependencies between the components. It is high in the case of efficient information pathways without redundancies. Note that both our measures of potential and connectedness are scaled with the total system throughput, i.e., the sum of all information transfers. The measures thereby increase with growing overall system activity.
Our definition of resilience is an analogue of the established spectral graph-theoretical measure of connectivity (Chung 1977). Connectivity quantifies the vulnerability of undirected graphs to perturbations. More precisely, it indicates how the loss of a single edge of the graph can be compensated via existing alternative pathways. Let us recall Gunderson and Holling’s definition of resilience as “the magnitude of disturbance that can be absorbed before the system changes the variables and processes that control behavior” (Gunderson and Holling 2002:28). In this sense, connectivity captures the idea of resilience as a measure of the “safety net” of the system’s information structure. We transferred the notion of connectivity to the case of directed graphs. See Appendix 1 for the mathematical definitions of the systemic variables.
The R package QtAC (Quantifying the Adaptive Cycle) allows for straight-forward application of our method (https://github.com/hannahschrenk/QtAC). The package is presented, and its application illustrated in Schrenk et al. (2022). The R code underlying the computations we used is provided in Appendices 2 and 3.
Data and parameters of the case study: Surtsey
Our analysis is based on a time series of vascular plant species’ abundance data collected in a 10 x 10 m plot (plot 1) on the southern part of Surtsey (see Fig. 1 b, d; Appendix 4). The plot is part of a gull nesting site on sand-filled sheet lava (Fig. 1 d). During the observation period, 12 different vascular plant species were apparent (see Table 1).
Data have been collected biennially in July by the Agricultural Research Institute and the Icelandic Institute of Natural History from 1990 to 2018. Next to species’ abundances, the number of gull nests within and around the plot was counted from 2004 onward. Because it is well-known that there are strong mutual effects between plants and breeding birds on the island, this quantity is included in our analysis in form of the variable nesting density (ND). For more information on data collection and the approach to monitoring, see Magnússon et al. (2009). Precipitation data have been obtained from the weather station on Stórhöfði, the main island in the Vestmannaeyjar Archipelago, about 18 km away from Surtsey (provided by the Icelandic Meteorological Office in January 2019, Appendix 5). We consider the 12 vascular plant species and the nesting density as components of a joint complex system. To estimate the information networks and to compute potential, connectedness, and resilience, we used the functions as implemented in the R package QtAC (https://github.com/hannahschrenk/QtAC).
Following the terminology in Appendix 1, we chose a fixed window size of w = 6, hence, the estimation of each information network is based on the 12 preceding years. The choice of this parameter is always a trade-off between statistical reliability (favored by a large number of values in the window) and a sufficient temporal resolution of the results. In general, we do not recommend using a window size of less than five data points (compare Schrenk et al. 2022). In the present case, there are two reasons for the choice of the relatively small window size of w = 6. First, with one data point corresponding to two years, the degree of averaging and thereby smoothing of the temporal resolution of the results strongly increases with the window size. Second, the larger the window size, the later the time point of the first estimated network. In this case study, we want to particularly observe the systemic effects of the arrival of the birds in 2004, hence, it is necessary to consider the system some time in advance.
We chose a uniform history length of k = l = 3. Because we interpolate the existing data for reasons of statistical reliability, this history length corresponds to a period of two years (compare Appendix 1). This choice is based on an assessment of the system dynamics. In regard to their temporally local nature, a history length of four years seems inappropriate. The data situation does not allow for a finer resolution.
An estimated transfer of information was only considered if its significance was ≤ 0.1. We do not recommend using a significance > 0.1 to guarantee statistical reliability of the estimations.
Resilience was normalized with respect to the maximal edge weight of the underlying network (compare Schrenk et al. 2022). Hence, the system’s resilience depends on the relative edge weights, only, rather than the total amount of information being transferred.
We set the amount of random Gaussian noise added during the estimation to 1e-20. This choice is based on a sensitivity analysis. Appendix 6 displays, for exemplary noise levels, mean and standard deviation of the systemic variables on basis of 50 runs each. Although potential and connectedness show little variation for noise levels below 1e-20 (Appendix 6 a, b), the standard deviation clearly increases for noise levels above this value (Appendix 6 c). In some years, the standard deviation of resilience remains comparably large even for small noise levels. This is due to the fact that resilience measures local vulnerabilities whereas potential and connectedness are global network properties. Thus, resilience is highly sensitive to the (non-) existence of single edges in the network. In such cases, it is advisable to search for reasons for the high standard deviation and to take these insights into account in the system analysis.
RESULTS AND DISCUSSION
To identify a representative development of the systemic variables, we consider the results for 50 runs of our method with the parameters set as specified in the Methods section. Figure 2 (a) displays the mean and standard deviation of these runs. In the case of both potential and connectedness, the standard deviation is small enough to identify the development of the respective systemic variable unambigously. However, the system’s resilience shows a comparably high standard deviation in 2000, 2002, 2006, 2010, and 2012. Figure 2 (b) displays the distribution of the values in the form of a scatterplot. One can observe that resilience is evenly distributed in 2002 whereas it is grouped around two values in the other years. The latter can be traced back to the (non-)existence of single edges, the significance of which is estimated to be close to the threshold level of 0.1. Therefore, the edge sometimes is included into the network while being excluded in other runs. We chose a value representing the majority of results as the resilience of the system. In the case of an even distribution (year 2002), we chose an average value as representative resilience. However, interpretation must be taken with care for this value.
Figure 1 (a, c) illustrates a representative development of the system’s systemic variables. The three-dimensional plot is shown from two different perspectives. The colors highlight the adaptive cycle phases identified, with green indicating phases of stability, red phases of decline, and orange phases of transition. Analyzing the community of vascular plants on the study plot reveals four clearly distinguishable phases.
Our observation period started with an r-phase from 2000 to 2004. Typically, potential and connectedness increased during this period. Resilience overall increased as well, which is rather unusual.
A clear K-phase can be identified from 2004 to 2006. Connectedness further increased, accompanied by a strong decline in resilience. Typically for the late K-phase, potential starts decreasing.
This phase is interrupted by a system breakdown starting in 2006. Because data were provided on a biennial basis, we decided to merge the subsequent two phases of stochasticity into an Ω/α-phase from 2006 to 2010. It starts with the characteristic decline of potential and connectedness along with a concurrent increase of resilience. From 2008 to 2010, potential and connectedness started increasing again while resilience still increased as well, which is typical for the α-phase.
The period from 2010 to 2012 revealed a second r-phase, indicated by increasing potential and connectedness, accompanied by a decrease in resilience. In 2012, the exploitation phase of the system was suddenly interrupted by a simultaneous decline in all three systemic variables. Following that year, the variables continued to oscillate around their current values. Their interplay did not exhibit the typical profile of one of the adaptive cycle’s phases. Such atypical development usually hints at external drivers, essentially changing the system’s characteristics.
Overall, the system ran through one complete cycle and started into a second one during the study period. From 2000 to 2006, a classical r/K-phase of exploitation and conservation preceded a four-year Ω/α-phase of release and reorganization. Although Gunderson and Holling anticipated the Ω/α-phase to be very short in comparison to the r/K-phase (Gunderson and Holling 2002), we conclude that in natural systems periods of reorientation and reorganization might take substantially longer.
In 2010, the system entered a new cycle, building up potential and connectedness until 2012. Interestingly, the expected further increase was interrupted in 2012. Such a deviation indicates changes in essential environmental conditions. Indeed, 2012 marked a special year of drought.
Appendix 7 (Fig. a) demonstrates that the overall pattern recurs under varying window sizes. Both for a window size of w = 5 and w = 7, the transition between two adaptive cycles during the study period is clearly recognizable. In particular, the sudden decline of potential and connectedness during the second exploitation phase is temporally stable. Note that the breakdown of connectedness during the first cycle shows a temporal shift from 2006 to 2010 with increasing window size. We will return to this phenomenon at a later time.
It is important to keep in mind that we cannot and should not expect any real system to exactly follow the idealistic pattern of the adaptive cycle. The metaphor should rather be seen as a description of the baseline development of a system that can be used as a background for a detailed analysis of a given instantiation. In particular, deviations from the ideal allow us to identify specificities in system development that are due to the particular interaction patterns given in a concrete situation.
The first r- and K-phase: the arrival of the birds
Let us first consider species composition during the period 2000-2004. Robust species being capable of surviving under harsh conditions still prevailed. Honckenya peploides and Leymus arenarius, along with Cerastium fontanum made up more than 80% of the land cover, and all of them were indicative of the earliest plant colonization on Surtsey (Magnússon et al. 2014). Toward the end of that period, the contribution of these early colonizers decreased to less than 50%. In 2004, the grass species Poa pratensis became dominant. This perennial grass forms dense mats and outcompeted annual community members such as Poa annua and Sagina procumbens. Plant community composition exhibited a clear transition from early pioneers toward a grassland community. The well-established association of Honckenya with Leymus, which has been important in establishing sustainable plant life on the island (Fridriksson 1992), started to give way to species of higher competition under stabilized environmental conditions. This development was associated with the increasing nutrient content of the soil, which can be traced back to the growing population of breeding seabirds on the island, bringing food for their chicks from the ocean to land (Magnusson et al. 2009). Two new colonizers settled down in the plot during this period: Festuca rubra in 2002 and Tripleurospermum maritima ssp. maritima (synonym Matricaria maritima) in 2004.
In the networks of information transfer, we gain insight into the inner functioning of the plant community (see Fig. 3). The arrival of the new species is clearly reflected in the network structure. In 2002, Tripleurospermum maritima ssp. maritima, previously peripheral, takes a central position in the information network (Fig. 3 a, b). Two years later, in 2004, the same phenomenon occurs with Festuca rubra (Fig. 3 c). Just like Leymus and Poa pratensis, F. rubra forms dense mats by extensive lateral spread above and below ground (Magnusson et al. 2009). In the same year, with the first gull nests being found in the plot (Fig. 4), nesting density moves into a central position in the network. Figure 5 (a, b) illustrates this sudden increase in information out- and inflow for these components. Although the information outflow of a species can be interpreted as the extent to which other species react to its development, we understand information inflow as an active process of adapting to the environment. In the case of the two new colonizers, information inflow dominates in 2004, reflecting their ‘active’ position upon arrival. Appendices 8 and 9 demonstrate the stability of the observed phenomena under varying window sizes.
Between 2004 and 2006, this trend in the community composition strengthens. Honckenya peploides, Cerastium fontanum, Stellaria media, and Poa annua further decrease in abundance, while Poa pratensis and Leymus arenarius strengthen their dominance. A plausible trigger for this change could be the accumulated effect of the gulls having been breeding in this area of the island for several years. The birds serve both as provider of nutrients (in particular nitrogen), as well as source of seeds (Magnusson et al. 2009, 2014). Thereby, they change the environmental conditions for all components of the system.
This intensification of the development of the previous years can be clearly observed in the system’s information network as well (Fig. 3 d). Tripleurospermum maritima ssp. maritima, Festuca rubra, and nesting density are closely connected, building a central point of the network. Figure 5 (b) shows that information inflow of Festuca rubra has further increased. Another central position in the network is taken by Taraxacum, which enters the plot in 2006. This is also reflected in its increase in information in- and outflow in 2006 in Figure 5 (d).
Compared to 2004, the overall structure of the information network has changed from many rather equally weighted edges to fewer, more heterogeneously weighted edges. This change in topology reflects the focusing that has taken place within the system structure and is responsible for the increase in connectedness and the decrease in potential. From an ecological point of view, the extremely low resilience at the end of this period can be further explained by still prevailing, rigid associations of the pioneers Honckenya and Leymus. These connections are now slowly dissolving with the decline of Honckenya.
The first breakdown: from pioneers to perennials
Between 2006 and 2010, there were no remarkable changes in community composition. The plot was dominated by the higher competitive species Poa pratensis and Leymus. Both plants together provided almost all of surface cover during this period (see Fig. 1 b). However, the preceding reorganization eventually resulted in a system breakdown. From 2006 to 2010, the network structure strongly changed (Fig. 6), being characteristic for the Ω/α-phase of a complex system. Trying to adapt to changing environmental conditions, the system exploits different opportunities in a “trial-and-error” mode. In 2008, Tripleurospermum maritima ssp. maritima was no longer part of the closely connected triangle with Festuca rubra and nesting density (Fig. 6 a). Instead, Taraxacum has developed close connections with the latter two. Figure 5 (a) demonstrates that meanwhile, the information outflow of nesting density dominates, capturing the strong influence of the breeding birds on the vegetation. This phenomenon can be explained by the fact that the specialized and demanding species of the second succession wave rely on the nutrient input by the birds. Furthermore, the effect of bird disturbance has to be considered, which might allow the new species to colonize and expand.
Recall the temporal shift of the system breakdown under varying window size (Appendix 7). This phenomenon hints toward the fact that certain causes for the breakdown can be traced back to the beginning of the time window. More precisely, we probably see late impacts of the fundamental restructurings in community composition following the year 1996. Hence, a similar pattern should be observable in the information transfer of certain species. As an example, consider how the sudden decline in information in- and outflow of Puccinellia distans is shifted from 2006 to 2010 with increasing window size (Appendix 10). Indeed, Puccinellia is one of the plants peaking in abundance in 1996 and declining from then on.
In 2010, the network stabilizes as indicated by a rather balanced set of edges with more equally distributed flows of entropy. This indicates the end of the Ω/α-phase and the start of a new period of growth. Considering the species’ information flow, future trends emerge. Although the total information flow of nesting density and Festuca rubra strongly decline, the information flow of Stellaria media starts increasing (Fig. 5 a-c). Furthermore, the information network reveals an increasingly central position of Tripleurospermum maritima ssp. maritima (Fig. 6 b). On a system level, the accumulated nutrients and the newly established plant colonizers led to an increase in potential between 2008 and 2010.
The radical restructuring in the information networks during 2006 and 2010 is particularly apparent in the development of the components’ eigencentralities (compare Csardi and Nepusz 2006). Simply put, in a network, nodes with high eigencentrality are strongly connected to other nodes that themselves are strongly connected too. Thus, eigencentrality captures the “importance” of a node within the network. Figure 7 shows the components’ eigencentralities in the information network between 2000 and 2018 in form of a heatmap. It is obvious that there is a turnaround between 2008 and 2010. Although eigencentrality of nesting density and Festuca rubra strongly declines (see also their sudden peripheral position in the network), eigencentrality of the grasses Poa annua and Puccinellia distans considerably increases. Furthermore, as already noticed in the information network, Stellaria media suddenly takes a central position.
The second cycle: the failure of a plan
Between 2010 and 2012, many species increased in abundance, among them Poa pratensis, Leymus arenarius, and Stellaria media. In the system’s information network of 2012, the development of the preceding two years has intensified (Fig. 6 c). Both Tripleurospermum maritima ssp. maritima and Stellaria media took central positions in the network. As well, Taraxacum gained a central position again. The information network shows the typical “contraction” behavior of a system’s r-phase, focusing on a few information transfers. This behavior results in an increase in connectedness and a decrease in resilience.
From 2012 to 2014, the system’s r-phase was suddenly interrupted by a simultaneous decline in all three systemic variables. The system composition shows a drastic change as well: almost all species are decreasing in abundance between 2012 and 2014 with Honckenya and Cerastium becoming extinct at the plot (Fig. 1 b). This phenomenon can be traced back to a lack of water perturbing the system during its move of exploitation. Figure 8 (a) shows the mean rainfall during the vegetation period as being measured at Vestmanneyjar weather station, 18 km away from the island of Surtsey. In 2012, precipitation was at a long-term minimum of less than 50 mm. Although the amount of precipitation was comparably low in 2007, the plants had never shown such severe drought symptoms before (B. Magnússon, personal communication, October 2020). Observe that the gross primary production of plot 1 (Appendix 11) displayed in Figure 8 (b) shows a minimum in 2012. One reason for this phenomenon might be the fact that in 2012, the period with no rain at all was longer than in 2007 and showers were lighter, unable to really wet the soil (B. D. Sigurdsson, personal communication, November 2020). In addition, a peak in the population of birds in 2012 (see Fig. 4 b) presumably provided additional stress for the plants. More precisely, high nutrient input led to a reduction of belowground growth, increasing the plants’ sensitivity to drought stress. Beyond these external reasons, we trace the extreme consequences back to the fact that the drought was hitting the plant community at the onset of its exploitation phase, whereas, in 2007, the system was in its Ω-phase.
Although the system has been shaped under humid climatic conditions and moderate disturbance through birds, the plant community was suddenly confronted with completely unexpected conditions. The ‘plan laid out’ during the α-phase was doomed to fail because of the unexpected drought. The community exhibited features of a system finding itself in conditions it had not been adapted to. Under these circumstances, the system is vulnerable, indicated by its extremely low resilience (Fig. 1 a). Following 2012, the system (temporarily) has to invest into managing extreme environmental conditions rather than building up potential for future growth. We note in passing that no data have been taken in odd numbered years. Thus, the exact tipping point of the system cannot be determined within a year’s precision.
Considering the information networks during this period, one can observe that Stellaria took a highly central position between 2012 and 2014 (Fig. 6 c, d). Figure 5 (c) further illustrates this fact and, moreover, emphasizes Stellaria’s active role in the community by a dominance of information inflow in 2016. This phenomenon is clearly visible in the respective information network (Fig. 9). The active role of Stellaria during this period of change can be ecologically underpinned. Note that it is Stellaria only that can increase during this year, profiting from released nutrients through dead plant material and by exploiting the opening of an unexpected opportunity (Fig. 1 b). Being a short-lived, annual plant of typically high growth rate and capable of producing seeds rapidly and abundantly (Grime et al. 1988), Stellaria profits from the opportunity being given by the environmental perturbation.
Synopsis on the adaptive cycle
The adaptive cycle metaphor has been the subject of debate since its initial publication. Questions arose concerning the interpretation of the three characteristic variables connectedness, potential, and resilience. There were also questions addressing the rigor of the four phases to be passed through in their given order. From a system science perspective, the identification of a complex system always includes the perspective of the observer. Thus, there can be no universal physical instantiation of the characteristic variables because their interpretation necessarily is subject to what pattern an observer considers as a system. However, following our understanding of a system, i.e., a set of interacting agents, allows for an interpretation of the variables in terms of characteristics of interaction. Thinking of an edge in a system’s information network as a communication channel, our measure of connectedness captures the unevenness of the distribution of communication channels. Connectedness rises once a system starts to make more use of some of the interactions while rendering others less important. Similarly, potential captures the average variability a system can make use of if encountering unprecedented conditions. Large potential thus implies that the system has degrees of freedom in its “communication structure.” Finally, resilience measures the susceptibility of the network to lose an interaction upon external perturbation.
The adaptive cycle metaphor captures the natural sequence of interrelations among these variables resulting from a system’s effort in maintaining viability. Deviations from this ideal being observed in the concrete analysis therefore enable us to learn more about a specific system in a similar way because theoretical concepts allow us to make sense of experimental observations. It is in this sense that the second cycle identified in the Surtsey data leads us to conclude that something changed substantially in the environment of the plant community. Likewise, the first complete cycle reflects the expected adaptation of the plant community to cope with the rising appearance of birds.
Consequently, it is not necessary to assume the phases of the adaptive cycle are of a certain relational duration. Although it appears to be convincing to expect the Ω/α-phase to be shorter than the r/K-phase, such a conclusion does not make sense from a system science perspective. The duration of the r/K-phase, for example, reflects the predictability of the environmental conditions as well as the overall stability. In the Surtsey study, it seems to take more time for the plant community in plot 1 to stabilize after the exceptional perturbation in 2012. In the end, adaptation of life requires and implies the presence of randomness, one of the main characteristics of the α-phase.
CONCLUSION
We provided a holistic analysis of the development of a vascular plant community on the volcanic island of Surtsey between 2000 and 2018. Based on our abstract approach of analyzing system development, a refined analysis reveals the role of certain plants as drivers of change under varying environmental conditions. We have been able to identify two adaptive cycles during the period of study. A first breakdown in 2006 can be traced back to the vast changes in community composition that the system experienced in the preceding years. Indeed, the network of information transfer reflects this shift from a pioneer to a classical grassland community. Our network analysis hereby backs the hypothesis that the boost of nutrients brought into the plant community by the breeding gulls is the main driver of this process of change. With the grass species having assured their dominant position in the plant community, another cycle starts around 2010. Atypically, the exploitation phase is interrupted by a sudden decline in all three variables. During the following years, the variables oscillate without exhibiting characteristics of a specific phase of the adaptive cycle. We trace this phenomenon back to a severe drought in 2012.
The development of the vascular plant community on Surtsey is fairly well understood. The individual species are well studied, as well as their interplay with each other and their coping with environmental factors. Therefore, the plant community on the island of Surtsey provided an ideal scenario to study suitability of the adaptive cycle metaphor in analyzing living systems. Comparing the development of a system with the idealistic scenario given by the adaptive cycle, deviations provide insight into drivers of internal dynamics. The case study demonstrates the capability of the method to analyze systems less studied and to reveal patterns of change. The approach, based on information networks, provides a basis to unravel the systems’ interaction structure and thereby the systemic function of individual components.
Because the approach can also be applied when comparing various systems, the database of observations of life on Surtsey provides further opportunities for better understanding the development of life on a newly formed island. Distributed over the island, there are several permanent monitoring plots exposed to different soil, weather, and environmental conditions. In particular, the level of influence by seabirds and seals strongly differs across the island (Magnússon et al. 2020). Comparing these different plant systems following the adaptive cycle metaphor might reveal new insights into adaptive strategies of plant communities in exploring new reservoirs of resources.
Aside from the ecological insights, this case study provides a valuable testing scenario for the possibilities and limits of our method. It turns out that a certain background knowledge of the system components and their interplay can be used for finding reasonable choices for the parameters. Nevertheless, parameter choices should be supported via sensitivity analyses.
Considering specific parameters, special attention should be paid to the level of random Gaussian noise added in the estimation of transfer entropy. An appropriate noise level is found if estimates lead to stable values of the systemic variables. However, the presented study shows that resilience, being a local measure, can nevertheless remain unstable. In such cases, causes should be identified, and the corresponding values of resilience should be interpreted with care. Another crucial parameter in the estimation process is given by the history length of a variable. Background knowledge of the corresponding component becomes particularly important. If such knowledge is not available, one can make use of established methods to estimate this parameter (see for example Lizier 2014). The choice of the window size is always a trade-off between statistical reliability and temporal resolution of the results. Having chosen a window size, we recommend verifying stability of the results under slightly varying window sizes.
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.DATA AVAILABILITY
The data that support the findings of this study are provided as supplementary material to this article. They include species' abundance data (Appendix 4), precipitation data (Appendix 5), as well as gross primary production data (Appendix 11). The R code used for generating the figures in the main body of the paper is provided in Appendix 2 and 3 (Appendix2.pdf, Appendix3.md). All functions used are provided in the R package QtAC (https://github.com/hannahschrenk/QtAC).
LITERATURE CITED
Bossel, H. 1992. Real-structure process description as the basis of understanding ecosystems and their development. Ecological Modelling 63(1):261-276. https://doi.org/10.1016/0304-3800(92)90072-M
Burkhard, B., B. D. Fath, and F. Müller. 2011. Adapting the adaptive cycle: hypotheses on the development of ecosystem properties and services. Ecological Modelling 222(16):2878-2890. https://doi.org/10.1016/j.ecolmodel.2011.05.016
Castell, W., and H. Schrenk. 2020. Computing the adaptive cycle. Scientific Reports 2020(10):18175. https://doi.org/10.1038/s41598-020-74888-y
Chung, F. R. K. 1997. Spectral graph theory. CBMS regional conference series in mathematics. American Mathematical Society, Providence, Rhode Island, USA.https://doi.org/10.1090/cbms/092
Cover, T. M., and J. A. Thomas. 2006. Elements of information theory. Wiley-Interscience, Hoboken, New Jersey, USA.
Csardi, G., and T. Nepusz. 2006. The igraph software package for complex network research. InterJournal. https://igraph.org
Fath, B. D., C. A. Dean, and H. Katzmair. 2015. Navigating the adaptive cycle: an approach to managing the resilience of social systems. Ecology and Society 20(2):24. https://doi.org/10.5751/ES-07467-200224
Fridriksson, S. 1992. Vascular plants on Surtsey 1981-1990. 1992. Surtsey Research Progress Report 10:17-30. [online] URL: https://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.535.2177&rep=rep1&type=pdf
Fridriksson, S. 2005. Surtsey. Ecosystems formed. University of Iceland Press,
Grime, J. P., J. G. Hodgson, and R. Hunt. 1988. Comparative plant ecology: a functional approach to common British species. Springer, Dordrecht, The Netherlands.
Gunderson, L. H., and C. S. Holling. 2002. Panarchy: understanding transformations in human and natural systems. Island, Washington, D.C., USA.
Holling, C. S. 2001. Understanding the complexity of economic, ecological, and social systems. Ecosystems 4(5):390-405. https://doi.org/10.1007/s10021-001-0101-5
Lizier, J. T. 2014. JIDT: an information-theoretic toolkit for studying the dynamics of complex system. Frontiers in Robotics and AI 1(2). https://doi.org/10.3389/frobt.2014.00011
Magnússon, B., G. A. Gudmundsson, S. Metúsalemsson, and S. M. Granquist. 2020. Seabirds and seals as drivers of plant succession on Surtsey. Surtsey Research 14:115-130. https://doi.org/10.33112/surtsey.14.10
Magnússon, B., S. H. Magnússon, and S. Fridriksson. 2009. Developments in plant colonization and succession on Surtsey during 1999-2008. Surtsey Research 12:57-76.
Magnússon, B., S. H. Magnússon, E. Ólafsson, and B. D. Sigurdsson. 2014. Plant colonization, succession and ecosystem development on Surtsey with reference to neighboring islands. Biogeosciences 11(19):5521-5537. https://doi.org/10.5194/bg-11-5521-2014
Odling-Smee, F. J., K. N. Laland, and M. W. Feldman. 1996. Niche construction. American Naturalist 147(4):641-648.
Odum, H. T. 1983. Systems ecology: an introduction. Wiley, New York, New York, USA.
Pianka, E. R. 1970. On r- and K-Selection. American Naturalist 104(940):592-597. https://doi.org/10.1086/282697
Schreiber, T. 2000. Measuring information transfer. Physical Review Letters 85(2):461-464. https://doi.org/10.1103/PhysRevLett.85.461
Schrenk, H., C. Garcia-Perez, N. Schreiber, and W. zu Castell. 2022. QtAC: an R-package for analyzing complex systems development in the framework of the adaptive cycle metaphor. Ecological Modelling 466:109860. https://doi.org/10.1016/j.ecolmodel.2021.109860
Ulanowicz, R. E. 1986. Growth and development: ecosystems phenomenology. Springer-Verlag, New York, New York, USA.
Ulanowicz, R. E., S. J. Goerner, B. Lietaer, and R. Gomez. 2009. Quantifying sustainability: resilience, efficiency and the return of information theory. Ecological Complexity 6(2):27-36. https://doi.org/10.1016/j.ecocom.2008.10.005
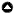
Fig. 1

Fig. 1. (a) Potential, connectedness, and resilience of the vascular plant system between 2000 and 2018. (b) Species’ abundances in plot 1 between 1990 and 2018. (c) Three-dimensional plot of the system’s course through the adaptive cycle between 2000 and 2018 from two perspectives. (d) Location of plot 1 on the island of Surtsey (edited version of Figure 2 in Magnússon et al. 2009).
Fig. 2

Fig. 2. (a) Potential, connectedness, and resilience of the vascular plant system between 2000 and 2018. (b) Species’ abundances in plot 1 between 1990 and 2018. (c) Three-dimensional plot of the system’s course through the adaptive cycle between 2000 and 2018 from two perspectives. (d) Location of plot 1 on the island of Surtsey (edited version of Figure 2 in Magnússon et al. 2009).
Fig. 3

Fig. 3. Information networks of the vascular plant community in (a) 2000, (b) 2002, (c) 2004, and (d) 2006. The width of an edge represents the strength of the information transfer between two components. Species abbreviations expanded in Table 1.
Fig. 4

Fig. 4. Number of nests recorded within and around plot 1 (area of 1000 m²) between 2000 and 2018. Note that nests have not been counted before 2004.
Fig. 5

Fig. 5. Information in- and outflow of (a) nesting density, (b) Festuca rubra, (c) Stellaria media, and (d) Taraxacum between 2000 and 2018. Information is given as nats (natural information unit).
Fig. 6

Fig. 6. Information networks of the vascular plant community in (a) 2008, (b) 2010, (c) 2012, and (d) 2014. The width of an edge represents the strength of the information transfer between two components. Species abbreviations expanded in Table 1.
Fig. 7

Fig. 7. Heatmap of the system components’ eigencentralities in the information network between 2000 and 2018. Species abbreviations expanded in Table 1.
Fig. 8

Fig. 8. (a) Mean rainfall (May-July) from 2000 to 2018 measured at the Vestmanneyar weather station. (b) Light saturated ecosystem photosynthesis rate (GPP) ± SE in plot 1 between 2006 and 2018. Measurements of GPP were done by an EGM-4 portable gas analyzer and with repeating flux measurements four times with a transparent and then a dark chamber. The value in 2016 is not a direct measurement but was derived from repeated measurements of the normalized difference vegetation index of plot 1.
Fig. 9

Fig. 9. Information networks of the vascular plant community in (a) 2016 and (b) 2018. The width of an edge represents the strength of the information transfer between two components. Species abbreviations expanded in Table 1.
Table 1
Table 1. Species of vascular plants recorded in Plot 1 between 1990 and 2018.
| Species | Abbreviations |
| Cerastium fontanum | CF |
| Cochlearia officinalis | CO |
| Festuca rubra | FR |
| Honckenya peploides | HP |
| Leymus arenarius | LA |
| Tripleurospermum maritima ssp. maritima (synonym Matricaria maritima) | MM |
| Poa annua | PA |
| Poa pratensis | PP |
| Puccinellia distans | PD |
| Sagina procumbens | SP |
| Stellaria media | SM |
| Taraxacum | T |